
Хранящийся в Эрмитаже скелет ребенка археологи нашли в конце 1920-х годов у села Мальта на территории современной Иркутской области. В том месте располагалась одна из самых известных позднепалеолитических стоянок Сибири. Четырехлетнего мальчика, погребли около Белой реки 24 000 лет назад. В ходе изучения ДНК ребенка ученые установили, что он был мальчиком примерно четырех лет. Генетики также выявили, что ДНК ребенка совпадает с ДНК современных североамериканских индейцев на 14-38%. Это объясняет тот факт, почему найденные в Америке останки предков современных индейцев имеют европейские черты. Кроме этого, соплеменники мальчика имели генетические связи с людьми, которые населяли европейскую часть России, а также современные Чехию и Германию.
Исследование позволяет пересмотреть время, когда предки индейцев пришли на Аляску, а затем и в Северную Америку. По-видимому, это произошло гораздо раньше, чем считалось (ранее 14,5 тысяч лет назад).
Результаты своих исследований антропологи и генетики опубликовали в журнале Nature (Raghavaan et al. 2013).
Происхождение первых американцев остается предметом споров. Хотя коренные американцы , кажется, генетически наиболее тесно связаны с восточными азиатами , среди ученных нет консенсуса в отношении того, какие именно группы населения Старого Света ближе всего к америндам. В работе Rahgavaan et al.2013 была рассмотрена последовательность генома останков доисторического человека ( МА- 1, датировка 24 000 лет ) со стоянки в Malta (юго-центральная Сибирь) , на средней глубине покрытия генома 1 х. Насколько известно, на сегодняшний день это старейший из существующих геномов анатомически современного человеческого генома. Кроме того, в работе были представлены результаты исследования другого древнего генома (с Афонтовой горы), но в силу плохого качества сиквенирования, эти исследования не столь интересы (другой южно-центрально-сибирский образец , Афонтова Гора- 2 датируется примерно в 17000 лет, он характеризуется аналогичными аутосомно- генетическими сигнатурами, что MA- 1 , и это означает что в течение последнего ледникового максимума в этом регионе не было существенной смены населения).
Митохондриальный геном MA-1 принадлежит к гаплогруппе U-1, широко представленной среди популяций верхнего палеолита и мезолита европейских охотников-собирателей. По мнению Германа Дзибеля, тщательный анализ митохондриального сиквенса MA-1 позволяет установить косвенные свидетельства в пользу архаичной предковой ретенции в митогеномных сайтах 13350 и 16399. Для крупных клад U были найдены следующие соответствия с геномами денисовцев и неандертальцев:
U: A12372 (D1, 2)
U1: C12879 (D1, 2), G13104 (D1, 2), A15148 (D1, 2, N), C16249 (D1, 2),
U6: C16172 (D1, 2),
U3: T150 (D1, 2, N),
U4: C16356 (D1, 2),
U9: A3834 (D1, 2)
U7: C152 (D1, 2, N, большинство людей), A14569 (D1, 2).
Учитывая, что здесь мы имеем дело с прямыми совпадениями с ДНК архаичных гомининов, обнаруженных в том же районе Южной Сибири, что и ДНК из Мальта, Дзибел делает интересные интерполяции в свете последних обсуждений статьи Малярчука о существовании «архаичных примесей» в мтДНК современного человека.

Y -хромосома МА -1 принадлежит в базальной по отношению к современным западным евразийцев ветви, восходя к точке конвергенции с собственно типичными коренными индейскими Y-хромосомными линиями. По мнению исследователей с Молгена, из имеющихся 41990 снипов на Y-хромосоме позитивными оказались 849, спорными 207, no-call 23622, а сам образец принадлежит к (R)P227+ (R)P280+ (R)P285+ (R1)P231- (R1)P234- (R1)P236- (R1)P245- (R1)P294- (R2)M479-. Образец MA-1 имеет «предковые значения аллеля» в 5 R cпецифических снипах.

Как Y- хромосома R, так и мтДНК-гаплогруппа U встречаются относительно редко к востоку от озера Байкал , которое здесь рассматривается как естественный природный барьер для генетического влияния западной Евразии в восточной Евразии. И действительно, оба эти гаплогруппы отсутствуют у коренных американцев , так что пока не ясно , каким образом коренные американцы ( которые принадлежат к Y -хромосомным гаплогрупп Q и С и мтДНК-гаплогруппам А, В, С, D, X ) связаны с палеолитическими сибиряками. Очевидным кандидатом для этой связи является Y- хромосомная гаплогруппа Р (общий предок Q и R ) .
Аналогичным образом, определенные свидетельства близкого (в эволюционном плане) родства современных западных евразийцев и современных коренных американцев мы находим в анализе аутосомMA- 1. Примечательно, что результаты анализа аутосом апостериорно опровергают априорную гипотезу о близком родстве MA-1 c популяциями Восточной Азии. Это говорит о том , что население, связанные с современными западными евразийцы в далеком прошлом (около 24 000 лет тому назад), вопреки традиционному мнению, были гораздо шире представлены в северно-восточной части Евразии.
Кроме того, по нашим оценкам, от 14 до 38% генов индейцев могли появится в результате вливания генов со стороны этого древнего населения . Это вливание ,вероятно, произошло уже после расхождения предков америндов с остальной частью предковой популяции , но еще до диверсификации индейских популяций в Новом Свете .
Ранее я проводил анализ MA-1 на предмет выявления процентного соотношения компонентов в программе Admixture. Мои вычисления можно резюмировать следующим образом:
Uralic (пик у селькупов и кетов — Западная Сибирь) 38,46%
Gedrosia-Caucasian (пик у брагуев и белуджей — Афганистан) 25,37%
North-Amerindian (пик у пима и апачей — юго-запад США) 17,38%
North-European-Baltic (пик у латышей) 11,5%
Ancestral-South-Indian 4,66%
Bantu 1,42%
Papuan-Australian 1,19%
Для сравнения внизу же приведены таблицы компонентов в образце MA-1 взятые из обсуждаемой статьи (Raghavaan et al. 2013). Разумеется, точного соответствия между компонентами нет, но в рамках произвольной аппроксимации можно предположить, что у MA-1 представлены «Меланезийский» (малиновый), «Америндский» (оранжевый) 16%, «Циркумполярный» (розовый), «Индийский» или «Гедрозия» (зеленый) 37%, «Восточноевропейский» (синий) 34%.

Самое интересное, что есть доказательства реальности обмена генов между образцом MA- 1 и коренными американцами. Данное доказательство имеет смысл , поскольку MA-1 является своего рода «прокси» для сибиряков периода, непосредственно предшествующего начальной колонизации Америки. Интересно и то, что МА -1 нельзя отнести к восточноевразийским популяциям , что следует из теста D-статистики ( Papuan , Han ; Sardinian , MA- 1). Результат этого теста не является статистически значимых, так как MA- 1 связаны с ханьцем не теснее, чем cовременные папуасы (обратное утверждение справедливо для современных коренных американцев ). Так что, похоже , что поток генов между MA -1 и коренных американцев был направлен от предков коренных американцов к MA -1 , а не в обратном направление.
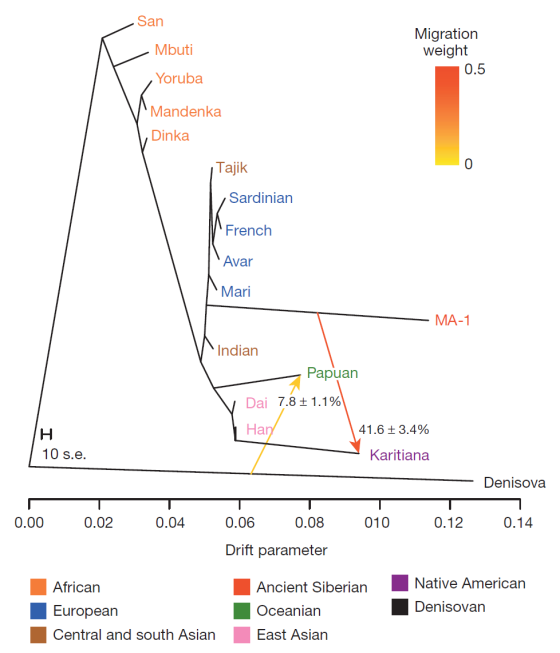
Поток генов от доноров вроде MA- 1 к реципиентам — предкам индейских предков мог бы объяснить, почему некоторые черепа первых американцев, были описаны как несущие в себя определенные морфологические характеристики , которые не напоминают аналогичные характеристики восточных азиатов.
Герман Дзибель, c отсылкой к работам известного уралиста В.Напольских (особенно «Древнейшие этапы происхождения народов уральской языковой семьи: данные мифологической реконструкции (прауральский космогонический миф)»), приводит еще одну интересную параллель, хотя она и не имеет прямого отношения к генетики. Мысль о том, что мифологические сюжеты и генетические маркеры должны между собой как-то коррелировать, для большинства ученных не нова: это слишком очевидно. А поскольку религии, в рамках которых существовали мифы, изначально были скорее родовыми, чем нацеленными на общечеловеческую экспансию, то и связь ген—миф должна быть, возможно, даже более тесной, чем ген—язык. В 1991 Напольских была предложена этноисторическая интерпретация распространения в Северной Евразии и Северной Америке космогонической мифологемы о создании суши из кусочка земли, принесённой ныряльщиком (животным или антропо- / теоморфным персонажем) со дна первичного океана (далее – миф о нырянии за землёй, в случае с нырянием птицы – миф о ныряющей птице, МНП). Было показано, что данный мотив имеет весьма древнее (как минимумверхнепалеолитическое) северноазиатское происхождение, и было обосновано предположение о том, что в наиболее древнем его прототипе (обозначенном кратко МНП0) сам образ ныряльщика (птица, млекопитающее, черепаха) на имел принципиального значения, а важнейшую смысловую нагрузку нёс успех последнего из нескольких ныряльщиков, который обеспечивался не физической, а особой сверхъестественной силой его. Сюжеты такого рода широко распространились в Северной Америке, и в большинстве продвинутых версий сохранили эту древнюю особенность, а конкретизация образа ныряльщика шла там в разных направлениях: в качестве такового могли выступать и птицы, и звери, и черепаха, и даже членистоногие (см. ниже). Согласно предложенной гипотезе, в Северной Евразии уже в глубокой древности получили распространение специализированные варианты мифа о нырянии за землёй с птицами в основной роли, в которых старая идея об успехе последнего и более слабого ныряльщика была со временем реализована в противостоянии птиц типа гагары, нырявшей первой и неуспешно, и утки (разные виды уток, распространённый вариант – поганка, очень редко – гусь или лебедь), принесшей землю. В более поздних работах Напольских (2011), Березкина (2012) и других был более точно определен ареал распространения мифологем. Этот ареал хорошо коррелирует с ареалом генетических маркеров.


Исследования международных групп ученых показывают, что люди достигли Америки более 30 000 лет назад — на 15 000 лет раньше, чем считалось ранее. Однако воздействие человека на вымерших крупных млекопитающих (мегафауна) произошло гораздо позже, когда популяции значительно увеличились. Эти результаты, опубликованные в журнале Nature, основаны на археологических исследованиях в пещере Чикиуите (Chiquihuite) в Центральной Мексике и статистическом моделировании дат из 42 археологических памятников.