
Автор этого произедения, Алексей Алексенко, несомненно, знаток биолгии и разных теорий её созаделей, то интересных, то идотических. Читая его работу с самого начала я долго возмущался как запутанно он излагает иногда несомненно умные мысли, а иногда полный бред. И он даже поясняет почему он построил свое повествование именно так, он по сложившийся традиции пытается выловить из моря околесицы, которую несет их коллега, какую-то действительно ценную идею, а всю чушь милосердно проигнорировать. Словословия в текстах не менее 90%. Очень хотелось иметь сокращенный текст, в котором были бы только факты. Но продвигась все дальше и дальше как-то проникся трудностями изложения темы – загадочности полового взаимодействия живых существ. Лишними стали представляться уже не всё 90% многословия. Перепост с сайта “Сноб” работы “Зачем живые любят друг друга” даже с сокращениями получился бы чудовищно громоздким, поэтому делаю конспект начала работы отражающий сущность полового разеления живых существ…
Надо сказать, что само название работы “Зачем живые любят друг друга” указывает на идею некой цели которой следуют живые существа, хотя кое где у автора проскакивает мысль, что цели вообще-то нет. Тем не менее в описании любого взаимодействия элементов жизни рефреном звучит этот вопрос: “Зачем это?”. Ответ на этот вопрос автор не дает, поясняя, что разных теорий, объясняющих существование секса измышленно очень много. (Реально, вопрос “зачем?” в эволюции бессмыслен. Когда-то что-то как-то получилось и оказалось жизнеспособным, так оно и сохраняется и накапливается, а все неудачное погибло…)
Автор и появление эукариот из архей, захвативших бактерии ставших митохондриями, отнес к взаимодействию партнёров в неком аналоге секса. То есть получается, что секс некое подобие питания. Понятное дело, в философском плане такое мнение понять можно… Множество частей работы с рассказами о вариантах полового и бесполового размножения разных грибов, мхов, водорослей и девних животных иллюстрирующих идею, что секс не очень то и нужен можно пропустить. Конечно всё это всё иллюстрирует самое интересное в биосфере Земли, – жизнь позвоночных, но очень призрачно. Впрочем, некоторые примеры из жизни примитивных эукариот нужны для понимания явдения сексуального взаимодействия позвоночных.
Вот пример иллюстрирующий прообраз полового взаимодействия. Возьмём два плесневых грибка — чтобы один, например, нуждался для роста в аминокислоте аргинине, а другой — в триптофане. Для наглядности пусть у первого будут желтые споры-конидии, а у второго — белые (и то и другое — мутации: в норме споры зеленые). Перемешайте споры и посейте на среду, в которой нету ни аргинина, ни триптофана. Вместо того чтобы смиренно умереть в отсутствие питания, грибки начнут образовывать странные колонии, покрытые пестрым ковром из белых и желтых спор. Это дикарионы — грибы, в клетках (гифах) которых перемешаны два типа ядер. Ядра поддерживают друг друга: в одних есть ген синтеза аргинина, в других — триптофана, а вместе они обладают всем необходимым. Это, конечно, паллиативная мера: и желтые, и белые споры по-прежнему содержат по одному ядру и на среде без аминокислот не прорастут. Но хотя бы у них будет шанс дождаться лучших времен.
Итак, слияние клеток может быть полезно. Большинству бактериальных клеток мешает сливаться клеточная стенка: это довольно жесткая оболочка или даже скорлупка кнаружи от клеточной мембраны, изолирующая бактерию от внешнего мира. Но клеточная стенка есть не у всех. Возможно, лишившись клеточной стенки, две бактерии могли бы слиться буквально сами собой — по неосторожности. Тем не менее считается, что ни у каких бактерий и архей слияние клеток не является и никогда не было частью их повседневной жизни. Впрочем, недавние исследования показали, что это, возможно, и не так.
У самых разных эукариот есть ген HAP2, который кодирует белок, необходимый для вхождения спермия в яйцеклетку или слияния гамет. Сначала его нашли у растений, а потом похожие гены/белки стали находить у других организмов. Кстати, с названиями генов часто бывает так, что сначала у разных существ их называют по-разному, а потом, когда выясняется, зачем этот ген нужен, все его родственники постепенно приобретают одинаковое имя (заимствованное обычно у того организма, где этот ген лучше изучен или впервые обнаружен). Чтобы не путать читателя, мы не будем каждый раз объяснять, что, например, название ZIP взято у дрожжей, HIM — у червей, а с(3)G — у мух. Так что знакомьтесь: HAP2 из растения арабидопсиса, он же резуховидка, присутствует у разных эукариот и необходим для объединения мужской и женской гамет. Откуда он такой взялся?
HAP2 кодирует белок, принадлежащий к семейству фузексинов (fusexins). Эти белки умеют соединять две клеточных мембраны и перетасовывать их компоненты таким образом, что образуется соединяющая две мембраны трубка. Фузексины встречаются даже у вирусов, облегчая вирусной частице проникновение через клеточную мембрану. Впрочем, вирусы могли позаимствовать свои фузексины от других организмов.
В этой истории попытался разобраться Бенджамин Подбилевич из Израильского института технологий в Хайфе вместе с коллегами из Уругвая и еще пяти стран, о чем они и написали статью. Сравнив известные фузексины, они установили, как бы мог выглядеть предковый белок, то есть ген. Оказалось, что очень похожие гены есть в геномах архей (тех самых архей, которые, приютив внутри себя бактерию, дали начало эукариотам, то есть нашей ветке жизни). Что делал фузексин внутри архей — действительно ли помогал им сливать свои клетки? Ну, по крайней мере мог бы. Чтобы доказать это, исследователи заставили этот реконструированный ген работать в культуре клеток млекопитающих, и эти клетки стали сливаться.
Хотя сами археи вроде бы сливаться не умеют, вполне возможно, что фузексин помогает им устроить нечто подобное половому процессу: к примеру, выстроить мостик от клетки к клетке, через который можно обменяться необходимыми генами. Но вот интересный поворот сюжета: у архей гены фузексинов находятся в геноме не где попало, а чаще всего в составе встроенных паразитических последовательностей, или «эгоистичных элементов», способных прыгать в хромосоме с места на место и размножаться быстрее, чем хозяйская клетка.
Если не обременять себя строгими доказательствами, вырисовывается заманчивая гипотеза. Изначально фузексин был частью вооружения эгоистичного элемента, генетического паразита, вируса. И это очень полезное оружие: без него элемент мог прыгать с места на место только в пределах одной клетки.
Как бы то ни было, но археи, оказывается, имели все необходимое для слияния клеток задолго до того, как их потомки-эукариоты воспользовались этим инструментарием, чтобы заняться сексом. Это открытие, кстати, часть большой драмы, которая прямо сейчас разворачивается в биологии. Долгое время казалось, что множество признаков сложных организмов не встречаются ни у бактерий, ни у архей — они как будто возникли на пустом месте после «великого симбиоза». Но постепенно один за другим появляются факты, свидетельствующие, что все это уже было — и было чаще всего именно у архей, очень плохо изученной, в отличие от бактерий, группы организмов. Конечно, большинство наших генов мы получили в наследство именно от бактерий — об этом подробно рассказано в книге Евгения Кунина «Логика случая». Но вот такие факты прямо-таки провоцируют меня по-дилетантски ляпнуть, что по существу все мы с вами, дорогие читатели, просто археи, сильно развившие свои природные дарования, но придумавшие не так уж много нового.
Вернёмся к двум сортам плесени Aspergillus nidulans, которые слили свои клетки, чтобы избежать голодной смерти. Одна плесень была с желтыми спорами и не умела сама синтезировать аминокислоту триптофан, а вторая — с белыми спорами и не могла синтезировать аргинин. Тем не менее, когда мы посадили их вместе на среду без аргинина и триптофана, у нас выросли колонии со смешанными бело-желтыми спорами — дикарионы, в которых после слияния клеток перемешались ядра двух наших грибков. Колонии росли на бедной среде, потому что два типа ядер могли совместно обеспечивать потребности гриба в аминокислотах. Однако ни белые, ни желтые споры-конидии не прорастали: в споре-то ядро только одно, а поодиночке они справляться так и не научились.
При этом, если смыть водой все-все споры с таких колоний и упрямо размазать их по бедной среде, кое-что все-таки вырастет. Кое-что не с белыми, не с желтыми, а с зелеными спорами, как у дикого гриба (и белые, и желтые споры — это мутанты). И эти споры будут заметно крупнее обычных, отчего колонии приобретают красивый чуть дымчатый оттенок. Кто это такой у нас тут вырос? А это диплоид! Где-то там, в глубине дикариона, два ядра слились в одно. Это не секс: не будем углубляться в тонкости, но слились они не так, как это делается при сексе, а просто между делом. В этих новых диплоидных ядрах каждая мутация из тех, что были у родителей, компенсирована здоровой копией гена в наборе, полученном от другого родителя. Поэтому нашему диплоиду не нужен для роста ни аргинин, ни триптофан, и споры у него вполне обычного для этой плесени зеленого цвета. Итак, для некоторых эукариот сливать ядра клеток — тоже не слишком хитрый фокус.
А теперь подвергнем нашего диплоида небольшой неопасной процедуре: посадим иголочкой несколько спор на питательную среду, куда добавлен вредный яд, мешающий хромосомам нормально расходиться при делении клеток. Сначала мы ничего не увидим: гриб просто не сможет расти, потому что при каждой попытке поделиться его ядра будут терять хромосомы. Но рано или поздно в какой-то части гриба все лишние хромосомы диплоида будут потеряны — он снова станет гаплоидом. И тогда мы увидим растущий сектор колонии плесени с зелеными, или белыми, или желтыми спорами. Возможно, там будут, к примеру, желтые сектора, нуждающиеся для роста в аргинине, но не нуждающиеся в триптофане.
И тут вы — если вы генетик — захлопаете в ладоши от радости. Дело в том, что мутации trpC, argC и yB находятся на одной (а именно, восьмой) хромосоме нашего гриба. И если у этих потомков, в отличие от родителя, совместились признаки «желтый» — «не нуждающийся в триптофане» — «нуждающийся в аргинине», это значит, что в восьмой хромосоме случились события рекомбинации. Гены родителей перетасовались между собой. При этом ни малейшего секса там не было и в помине. Не было и мейоза.
Это в 1954 году открыл итальянский генетик Гвидо Понтекорво, работавший в шотландском городе Глазго. То, что он открыл, назвали «парасексуальным процессом» — это такой секс без секса, то есть рекомбинация в неполовых, соматических клетках.
Собственно, с помощью таких опытов в середине ХХ века и было выяснено, как происходит рекомбинация. На мой взгляд, это одна из самых завораживающих страниц истории биологии: от разноцветных спор, бактериальных колоний и фаговых бляшек (это когда вирус нападает на бактерию, бактерии умирают, и на сплошном бактериальном ковре возникает прозрачное пятнышко) биологи протянули логическую нить к молекулярным перестройкам ДНК. И, как оказалось, все угадали правильно, хоть и не с первого раза.
Рекомбинация, как выяснилось, бывает не только у высших организмов, но и у бактерий, и у вирусов. Бактерии размножаются делением, то есть клонально и без всякого секса, но они обзавелись особыми приемами, чтобы обмениваться генами. Таких приемов, вообще говоря, три.
Первый называется трансформация. Бактерия умеет захватывать ДНК, плавающую во внешней среде, и потом встраивать ее в свой геном. Она делает это не по небрежности, а намеренно: у нее есть для этого специальные приспособления, которые запускаются в периоды стресса, например голодания. Если бактерии голодно, значит, возможностей ее генома не хватает на то, чтобы прокормить себя. Возможно, мешают какие-то мутации или просто нужного гена у нее никогда и не было. И тут микроб делает последнюю ставку: а вдруг нужный ген плавает где-то рядом? Это не так уж маловероятно, как кажется, потому что если бактерии плохо, то плохо и окружающим ее сестрам-бактериям, и многие из них наверняка уже погибли, их ДНК вытекла наружу и где-то тут плавает.
Именно генетическая трансформация бактерий стала решающим доказательством того, мягко говоря, немаловажного факта, что в основе жизни на Земле лежит ДНК. Это открытие было сделано в 1944 году, и Освальд Эйвери остался в истории как пример ученого, совершившего эпохальный прорыв в своей области, но не удостоившегося Нобелевской премии. Номинирован на нее он был несколько раз, в том числе в 1949 году, когда премию присудили за лечение психических заболеваний с помощью лоботомии. Вот такой неприятный случай в истории Нобелевского комитета.
Впрочем, перейдем к другим приемам, с помощью которых бактерии обмениваются генами. Второй способ называется трансдукция. За него отвечают вирусы-бактериофаги, которые иногда встраивают свой геном в хромосому бактерии, живут там некоторое время, а потом вырезают сами себя и начинают размножаться. Так вот, при вырезании они иногда могут по неосторожности прихватить кусочек бактериальной хромосомы с каким-нибудь геном. А после такой вирус заразит другую клетку, встроится в ее геном и подарит ей новый ген. Одно время было принято думать, что трансдукция — чистая случайность, и бактерии в ней уготована лишь пассивная роль. Но сейчас выясняется, что особые приспособления — «агенты переноса генов», или АПГ — являются частью бактериального аппарата выживания и эволюции, и не всегда легко бывает определить, где там злой вирус, а где добрый механизм клетки-хозяина. Об этом хорошо прочитать в «Логике случая» у Евгения Кунина, благо по самой первой биологической специальности он именно вирусолог и лишь потом стал биоинформатиком-теоретиком. Для наших скромных штудий эта тема — уже немного чересчур.
Наконец, есть третий способ, очень изощренный, и он называется конъюгация. О ней мы немного говорили: бактериальные клетки соединяются мостиком из цитоплазмы, и по этому мостику от одной клетки к другой передается особый генетический элемент, F-фактор. Элемент часто передается сам по себе, и тогда это похоже на обычную инфекцию. Но бывает, что элемент встроен в хромосому, и тогда он начинает перетаскивать всю хромосому за собой (а другая копия остается внутри клетки, так что никто не умирает). Весь процесс занимает около полутора часов, но, если встряхнуть колбу с конъюгирующими бактериями, он прервется раньше, и от донора к реципиенту перейдет лишь часть генов. А поскольку генетики знают, в каком именно месте хромосомы встроен F-фактор у их домашних питомцев-бактерий, они могут использовать этот фокус, чтобы составлять карты генов. Генетическая карта кишечной палочки размечена в минутах. Если какой-то ген успевает перейти в клетку-реципиент за 30 минут конъюгации (а за 29 не успевает) — значит, этот ген расположен «на 30-й минуте генетической карты».
Конъюгация уже так похожа на секс, что есть соблазн думать, будто это он и есть. У инфузорий — сложных ядерных организмов — половой процесс тоже называется конъюгацией, потому что внешне похож на бактериальный: две клетки соединяются мостиком и через него обмениваются ядрами. Но у инфузорий эти ядра — продукт мейоза, почти что наши гаметы, и все это просто часть обычного полового процесса. К тому же все происходит симметрично, родители вносят одинаковый генетический вклад. А при бактериальной конъюгации есть ярко выраженные «мама» и «папа» («папой» будем считать штамм, в котором был F-фактор и который инициировал все это безобразие). По окончании процесса «папа» остается с чем был, а «мама» может использовать «папину» хромосому, чтобы заменить ею кусочек своей. А может и не использовать. Так что это не совсем одно и то же.
Переходим к настоящему половому процессу.
В 1890 году благодаря Августу Вейсману, мтало известно, что число хромосом при оплодотворении удваивается, а затем при мейозе вновь возвращается к прежнему значению. Лишь в 1905 году появились термины «гаплоид» и «диплоид», и тогда же в употребление вошло слово «мейоз» — в переводе с греческого «уменьшение».
Мейоз начинается так: сперва все хромосомы в клетке удваиваются.
Здесь нам сразу придется сделать небольшую паузу, чтобы прояснить эту историю с «удвоением»: уж больно много всего там двойного, и некоторые с непривычки в этом путаются. Речь у нас здесь идет о диплоидной клетке. Это значит, что каждая хромосома в ней представлена в двух копиях — точнее, в двух вариантах, полученных от двух родителей и отличающихся друг от друга какими-то мутациями. Но, если посмотреть на такую клетку в микроскоп, этих интимных отношений между родительскими гомологичными хромосомами вы никак не заметите: обычно они не проявляют ни малейшей тяги друг к другу.
Далее: на картинках хромосомы часто предстают в виде этаких букв Х: две половинки связаны друг с другом в одной точке, которую называют центромерой. Эти половинки называют «сестринскими хроматидами». Вот они-то очень даже тяготеют друг к другу: появляются перед делением клетки в виде «иксов», соединенные центромерой, а после деления половинки иксов расходятся в разные стороны, то есть хромосомы становятся одинарными, по одной хроматиде в каждой. Одна хроматида — это одна молекула ДНК. Итак, перед клеточным делением у нас есть две гомологичных хромосомы, и, стало быть, четыре очень похожих друг на дружку молекулы ДНК (хроматиды). Идентичные хроматиды одной хромосомы называются «сестринскими», а хроматиды от хромосом папы и мамы будем называть просто гомологичными.
Наконец, каждая молекула ДНК — это двойная спираль, состоящая из двух нитей. В принципе, одна нить содержит всю генетическую информацию, какая нужна организму. Таким образом, в двух гомологичных хромосомах и четырех хроматидах эта информация повторена 8 раз.
Вот теперь можно продолжать. Итак, хромосомы удвоились — так же, как они это делают перед обычным клеточным делением, то есть митозом. Но вместо того, чтобы разделиться на хроматиды, которые при обычном делении расходятся по полюсам, во время мейоза происходит нечто другое: гомологичные хромосомы находят друг друга и соединяются вместе. Тут-то в них и происходит таинственная рекомбинация — перетасовка отдельных частей. Заметим, что в процессе участвуют вообще-то не две, а целых четыре молекулы ДНК, хотя рекомбинация между сестринскими хроматидами обычно не приводит ни к каким видимым последствиям.
Когда дело сделано, клетке надо поделиться. При обычном делении к каждой центромере прикрепились бы по две нити веретена деления, идущие с двух полюсов клетки, и они растащили бы хроматиды по своим углам. Но при первом делении мейоза к каждой центромере прикрепляются микротрубочки только с одной стороны. Они немного подтягивают гомологичные хромосомы каждую в свою сторону, и тут становятся видны те места, где произошла рекомбинация. Они называются «хиазмы»: там две гомологичные хромосомы (точнее, хроматиды) продолжают цепляться друг за дружку. Эти зацепки очень нужны, чтобы хромосомы правильно разошлись: когда их нет, вся конструкция не может «натянуться», и хромосомы нередко распределяются неправильно: в одной клетке лишняя, в другой не хватает. Это называется «анэуплоидия», из-за которой, к примеру, у некоторых людей наблюдается синдром Дауна.
Наконец, хиазмы расцепляются и гомологичные хромосомы расходятся восвояси. Теперь в каждой дочерней клетке их вдвое меньше, чем было до деления, — гаплоидный набор. Однако каждая хромосома по-прежнему состоит из двух хроматид, и теперь они не такие уж и «сестринские»: в них произошли разные события рекомбинации, а потому они отличаются друг от друга немногим меньше, чем гомологичные хромосомы до начала всей этой свистопляски. Теперь хроматидам надо еще раз разойтись по разным клеткам, и это происходит как при обычном клеточном делении — митозе — с единственной разницей: удваивать ДНК уже не надо.
Вот как этот классический мейоз выглядит на схеме:
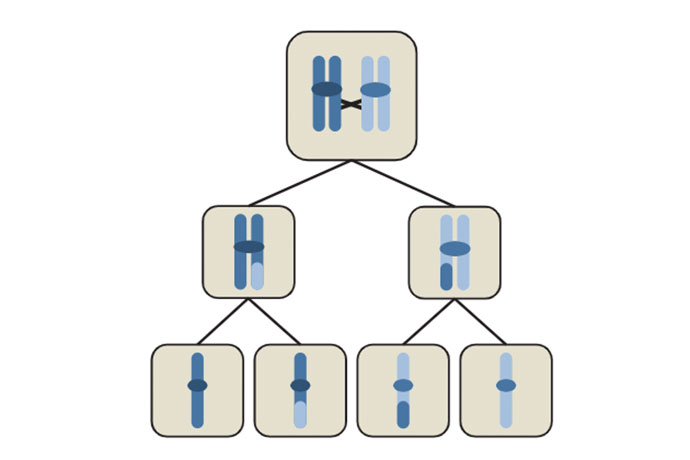
Итак, мейоз состоит из двух делений клетки — первое называют редукционным, второе эквационным, — в результате чего клетка из диплоидной превращается в гаплоидную. Вернее, в четыре гаплоидных. У сложных и продвинутых организмов вроде нас с вами или из этих четырех в яйцеклетку превратится только одна, а остальные становятся «полярными телами» и не участвуют в передаче генов потомству. Но вот у грибов такой дискриминации нет. Четыре клетки, получившиеся в результате мейоза, превращаются в четыре споры, или даже в восемь спор, потому что каждая проходит через еще одно деление.
В следующих разделах мы рассмотрим отдельные стадии мейоза и убедимся, как логично и естественно они выглядят, если предположить, что это не какой-то эволюционный скачок, а довольно экономичная адаптация того, что клетки и так уже имели, — обычного аппарата деления. А начнем, наверное, с рекомбинации.
В 1983 году вышла статья, в которой была представлена модель рекомбинации, более или менее соответствующая действительности. В этой модели разрыв ДНК был двойной! Молекула просто рвалась на два куска, да еще и эти куски сразу начинал подъедать с концов специальный фермент. В результате в той молекуле ДНК, которая выступала зачинщиком рекомбинации, появлялась огромная дыра, а длинный одноцепочечный хвост, который фермент оставлял несъеденным, начинал метаться по клетке, встревать в середину других ДНК и проверять, нет ли там гомологии. А когда она была — происходили последующие события, в том числе и образование структуры Холлидея. Вот схема этого процесса. Она называется «Модель починки двойного разрыва».
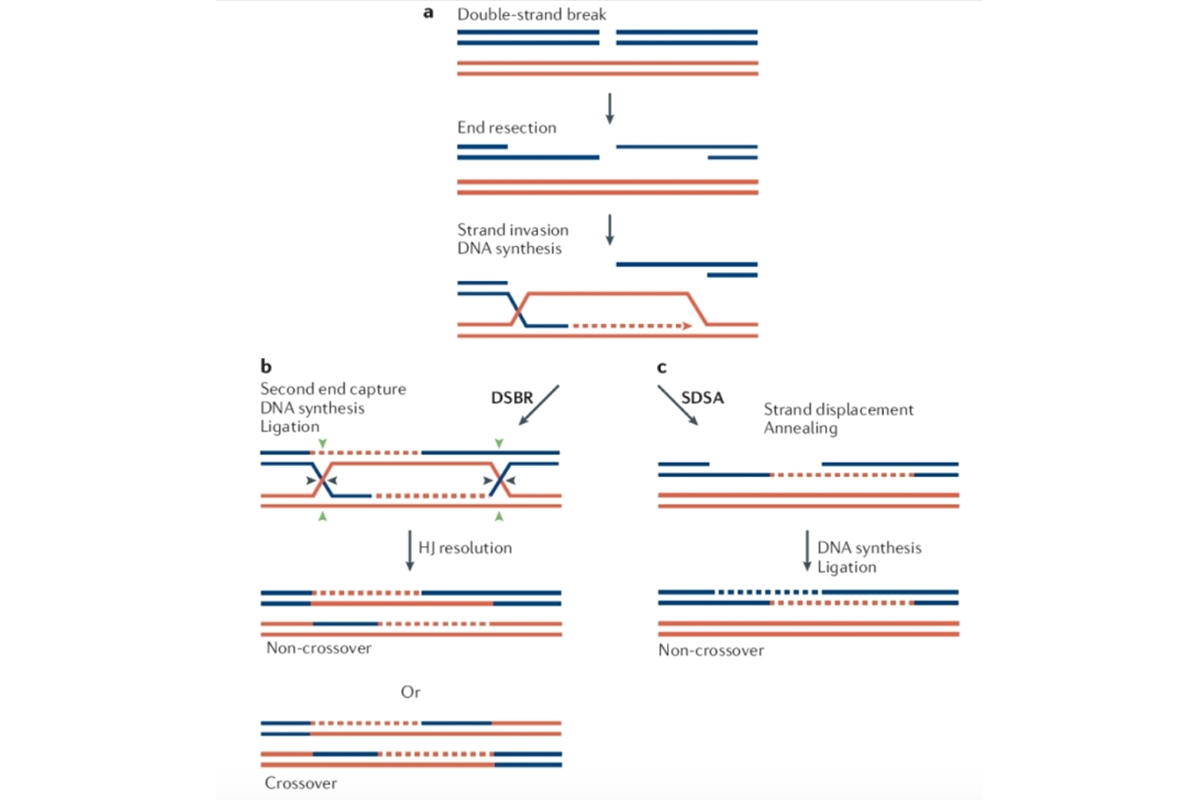
При одной конфигурации молекулы ДНК меняются флангами, то есть рекомбинируют все гены, лежащие справа и слева от точки разрыва. В альтернативном варианте изменения касаются только небольшой области неподалеку от точки разрыва, а фланги остаются на месте.
Во-вторых, те цепочки ДНК, которые получаются на выходе, на некоторых отрезках полностью покрашены в красный цвет, а кое-где красными оказываются три цепи из двух. Это и есть объяснение всех разнообразных вариантов генной конверсии, которые наблюдались в тетрадах грибных спор: вблизи от точки кроссинговера часть генетической информации одного из партнеров теряется.
В-третьих, на картинке видно, что теряются всегда гены синей молекулы, которая и затеяла весь процесс, причем она вступает в него уже изрядно потрепанной — с большой дыркой посередине. Зачем ей это нужно? Последовательности ДНК не любят теряться, их за это отбраковывает отбор. Однако игра стоит свеч, если в синей молекуле изначально был дефект: например, одна из букв-оснований оказалась подпорченной и ее пришлось вырезать, как раз с образованием двойного разрыва. Единственный вариант спасения — залатать дырку по образцу похожей молекулы.
А с чего бы возник этот дефект? Ну, например, под воздействием радиации.
Итак, одноцепочечный хвост изуродованной молекулы ДНК мечется по клетке в поисках партнера. Ему помогает в этом некий белок, который облепляет этот самый хвост со всех сторон. Это и есть тот самый белок, который у бактерии кишечной палочки называется RecA и который наш преподаватель иначе как «таинственным» не называл. Rec — это от слова «рекомбинация»: бактерии-мутанты по гену этого белка не умеют заниматься рекомбинацией, даже когда все остальные компоненты на месте. Очень похожий белок обнаружили и у высших организмов (сначала дрожжей, а потом и прочих, включая нас с вами), и называется он Rad51. Rad — это уже от слова «радиация»: мутанты по гену этого белка оказываются к ней очень чувствительны, потому что не умеют залатывать повреждения своей ДНК. Есть он и у архей, и там он тоже называется Rad. Это значит, что механизм рекомбинации, о котором мы сейчас толкуем, немыслимо древний и фундаментальный: им пользуются абсолютно все ветви жизни на Земле.
И еще один вывод, скорее исторического свойства. Как видим, иногда Rec=Rad, то есть механизмы рекомбинации и устойчивость организмов к радиоактивному излучению имеют много общего. Белку Rad51 (он же таинственный RecA) у людей помогают два интересных белка, Brca1 и Brca2. Они тоже в нужный момент оказываются там, на отчаянно ищущем помощи одноцепочечном хвосте ДНК. Их названия происходят от слов Breast Cancer — рак груди, — потому что мутации в соответствующих генах сильно повышают вероятность этой напасти. Отметим для себя, что раз небольшие дефекты этих белков чреваты столь серьезными последствиями, значит, рекомбинация не игрушка, и живому организму жизненно важно, чтобы там все прошло правильно. Причем важно не в контексте секса и размножения, а для повседневной жизни обычного соматического органа, хотя и очень красивого и косвенно связанного с сексом*.
Напрашивается фундаментальный вопрос, который останется без ответа. А может, секс — не главное в жизни? Ой, нет, спросим по-другому: может быть, рекомбинация, перетасовывающая гены, в том числе во время мейоза у высших организмов, возникла совсем не за этим? Может быть, это древний механизм ремонта повреждений, который гораздо позже был привлечен к новой работе: создавать разные комбинации генов и избавляться от мутационного груза (впрочем, мутационный груз и сам имеет прямое отношение к повреждениям ДНК)? Но именно из него, кажется, вырос весь этот масштабный проект под названием «мейоз» и «секс». А возможно, ради этого он и был затеян.