
Вагиш М. Нарасимхан *1,, Ник Паттерсон *3,2,, Прия Мурджани +5,4,, Nadin Rohland 21,, Ребекка Бернардос 1, Свапан Маллик 62,1, Иосиф Лазаридис 1, Натан Накацука 71,, Iñigo Olalde 1, Марк Липсон 1, Александр М. Ким 81,, Luca M Olivieri 9, Alfredo Coppa 10, Massimo Vidale 119,, Джеймс Мэллори 12, Вячеслав Моисеев 13, Егор Китов 1615,14,, Джанет Монж 17 лет, Николь Адамски 61,, Нил Алекс 18, Насрин Брумандхошбахт ‡6,1,, Франческа Кандилио 2019,, Кимберли Каллан 61,, Оливия Черонет 2221,19,, Брендан Дж. Каллетон 2423,, Мэтью Ферри 61,, Даниэль Фернандеш 2522,21,19,, Беатрис Гамарра 2621,19, Дэниел Гаудио 2119,, Mateja Hajdinjak 27, Éadaoin Harney 286,1,, Томас К. Харпер 2423,, Дениз Китинг 19 лет, Энн Мари Лоусон 61,, Мэтью Ма 62,1,, Кирстен Мандл 22 года, Меган Мишель ‡6,1,, Mario Novak 2919,, Jonas Oppenheimer ‡6,1,, Нирадж Рай 3130,, Кендра Сирак 3219,1,, Вивиан Слон 27 лет, Кристин Стюардсон 61,, Fatma Zalzala 61,, Чжао Чжан 1, Газиз Ахатову 15, Анатолий Н. Багашев 33 года, Алессандра Баньера 9, Баурыжан Байтанаев 15, Julio Bendezu-Sarmiento 34, Арман А. Бисембаев 3515,, Gian Luca Bonora 36, Темирлан Таргынов 37, Татьяна Чикишева 38, Петр Дашовский 39, Анатолий Деревянко 38, Miroslav Dobeš 40, Катерина Дука 4241,, Надежда Дубова 14, Мейрам Н Дуйсенгали 35, Дмитрий Эньшин 33, Андрей Епимахов 4443,, Suzanne Freilich 22, Алексей В Фрибус 45, Дориан Фуллер 46 лет, Александр Горячев 33 года, Андрей Громов 13, Сергей П. Грушин 47, Брайан Хэнкс 48 лет, Маргарет Джадд 48, Эрлан Казизов 15, Александр Хохлов 49, Александр П. Крыгин 50, Елена Куприянова 51, Павел Кузнецов 49, Donata Luiselli 52, Farhod Maksudov 53, Аслан М. Мамедов 54, Талгат Б. Мамиров 15, Кристофер Мейклджон 55, Дебора Си Мерретт 56, Roberto Micheli 579,, Олег Мочалов 49, Самариддин Мустафокулов 5853,, Аюши Наяк 41, Davide Pettener 59, Ричард Поттс 60, Дмитрий Ражев 33, Марина Рыкун 61, Стефания Сарно 59, Савенкова Татьяна Михайловна 62, Кулян Сихымбаева 63, Сергей М Слепченко 33, Ороз А. Солтобаев 37, Надежда Степанова 38 лет, Светлана Святко 6413,, Кубатбек Табалдиев 65, Maria Teschler-Nicola 6622,, Тишкин Алексей А 67, Виталий Викторович Ткачев 68, Сергей Васильев 6914,, Петр Велеминский 70, Дмитрий Воякин 7115,, Антонина Ермолаева 15, Мухаммад Захир 7241,, Валерий С. Зубков 73, Алиса Зубова 13, Синдэ Васанта 74, Карлес Лалуэса-Фокс 75, Matthias Meyer 27 лет, Дэвид Энтони 76, Николь Буавен +41,, Кумарасами Тангарадж +30,, Дуглас Дж. Кеннетт +‡77,24,23,, Майкл Фрачетти +79,78,, Рон Пинхаси +22,19,, David Reich 1,2,6,80,+
- Информация об авторе
- Информация об авторских правах и лицензии
PMCID: PMC6822619 NIHMSID: NIHMS1053677 PMID: 31488661
Версия этой статьи, подготовленная издателем, доступна на Science
Абстрактный
Секвенировав геномы 523 древних людей, мы показали, что основным источником происхождения современных жителей Южной Азии является доисторический генетический градиент между людьми, родственными ранним охотникам-собирателям Ирана и Юго-Восточной Азии. После упадка цивилизации долины Инда они смешались с жителями Юго-Восточной Азии и сформировали одну из двух основных групп предков жителей Южной Азии, чьи прямые потомки живут на юге Индии. В то же время они смешивались с потомками степных скотоводов, которые распространились по Центральной Азии после 4000 лет назад и сформировали другую основную группу предков. Степное происхождение в Южной Азии имеет те же черты, что и в Восточной Европе бронзового века, что указывает на перемещение людей, которое затронуло оба региона и, вероятно, распространило уникальные общие черты между индоиранскими и балто-славянскими языками.
Графический Реферат
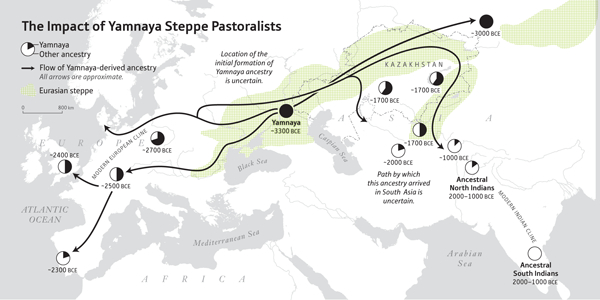
В бронзовом веке степные скотоводы ямной культуры распространились на два субконтинента: в Европу и Южную Азию. Круговые диаграммы отражают долю предков ямной культуры, а даты отражают самую раннюю доступную древнюю ДНК с предками ямной культуры в каждом регионе. Для ANI и ASI древняя ДНК пока не найдена, поэтому их диапазон определяется статистически.
Краткое содержание в одном предложении:
Полногеномное исследование древней ДНК 523 древних людей проливает свет на генетические обмены между степью, Ираном и Южной Азией и подчёркивает параллельные демографические истории двух субконтинентов: Европы и Южной Азии.
Краткое изложение на одной странице
Введение и обоснование:
Чтобы выяснить, в какой степени основные культурные преобразования, связанные с земледелием, скотоводством и изменениями в распространении языков в Евразии, сопровождались перемещением людей, мы представляем данные о древней ДНК 523 человек, живших в последние 8000 лет в основном в Центральной Азии и на севере Южной Азии.
Результаты:
Перемещение людей после появления земледелия привело к генетическим градиентам в Евразии, которые можно смоделировать как смешение семи сильно различающихся популяций. Ключевой градиент сформировался в Юго-Западной Азии в эпоху неолита и сохранился в бронзовом веке. На западе преобладали предки, связанные с анатолийскими земледельцами, а на востоке — предки, связанные с иранскими земледельцами. Этот градиент распространился на пустынные оазисы Центральной Азии и стал основным источником происхождения народов Бактрийско-Маргианского археологического комплекса бронзового века (БМАК). Это подтверждает идею о том, что археологически задокументированное расселение одомашненных животных сопровождалось распространением людей из нескольких центров одомашнивания.
Основное население BMAC не имело степных пастушеских корней и не внесло существенного вклада в формирование более поздних южноазиатских народов. Однако степные пастушеские корни появились у отдельных представителей BMAC на рубеже второго тысячелетия до н. э., примерно в то же время, когда они появились в южных степях. Используя данные о древних людях из долины Сват на севере Южной Азии, мы показываем, что степная раса распространилась дальше на юг в первой половине второго тысячелетия до н. э., составив до 30% предков современных групп в Южной Азии. Степная раса в Южной Азии имеет тот же профиль, что и в Восточной Европе бронзового века, что указывает на перемещение людей, которое затронуло оба региона и, вероятно, распространило уникальные общие черты между индоиранскими и балто-славянскими языками.
Основная предковая популяция современных жителей Южной Азии представляет собой смесь людей, связанных с ранними голоценовыми популяциями Ирана и Южной Азии, которую мы обнаруживаем у отдельных представителей двух культур, контактировавших с цивилизацией долины Инда (IVC), что позволяет предположить, что она была характерна для IVC. После упадка Индской цивилизации это население смешалось с северо-западными группами степного происхождения, образовав «древних северных индийцев» (ANI), и с юго-восточными группами, образовав «древних южных индийцев» (ASI), чьи прямые потомки сегодня живут в племенах на юге Индии. Смешение этих двух групп, возникших после Индской цивилизации, — ANI и ASI — сегодня определяет основной градиент генетической изменчивости в Южной Азии.
Заключение:
В более ранних работах было зафиксировано массовое переселение населения из степей в Европу в начале 3го тысячелетия до н. э., что, вероятно, привело к распространению индоевропейских языков. Мы выявили параллельную серию событий, приведших к распространению степной генетики в Южной Азии, тем самым задокументировав перемещения людей, которые, вероятно, способствовали распространению индоевропейских языков.
Введение
За последние десять тысяч лет произошли глубокие экономические изменения, вызванные переходом от собирательства к производству продуктов питания, а также произошли кардинальные изменения в культурных практиках, о чём свидетельствуют археологические находки, распространение языков и письменность. Степень, в которой эти изменения были связаны с перемещениями людей, оставалась загадкой для Центральной и Южной Азии отчасти из-за недостатка древних ДНК. Мы сообщаем о полногеномных данных 523 человек из Центральной Азии и самой северной части Южной Азии, начиная с эпохи мезолита (1), и анализируем их совместно с ранее опубликованными данными о древней ДНК со всей Евразии и данными о современных людях.
В Центральной Азии мы изучали, в какой степени распространение земледелия и скотоводства с Иранского нагорья на пустынные оазисы к югу от Степи сопровождалось перемещением людей или заимствованием идей у соседних групп (2–4). Что касается городских общин Бактрийско-Маргианского археологического комплекса (БМАК) бронзового века, мы оценивали, были ли люди, похороненные на его кладбищах, прямыми потомками более ранних мелких производителей продуктов питания, а также документировали их генетическую неоднородность (5). Далее на север и восток мы показали, что распространение сельскохозяйственных культур и одомашненных животных в эпоху ранней бронзы между Юго-Западной Азией и Восточной Евразией вдоль Внутреннеазиатского горного коридора (6) сопровождалось перемещением людей. Наконец, мы изучили, когда потомки ямной культуры, распространившейся по Евразийской степи примерно в 3300 году до н. э. (7–9), начали появляться в Центральной Азии к югу от степи.
В самой северной части Южной Азии мы сообщаем о временном отрезке, охватывающем более ста человек, начиная примерно с 1200 г. до н. э., который мы анализируем вместе с современными данными о сотнях современных южноазиатских групп, а также древней ДНК из соседних регионов (10). Согласно предыдущим исследованиям, большинство современных жителей Южной Азии относятся к генетической линии (11), которая, по-видимому, возникла в результате смешения двух сильно различающихся популяций около 4000 лет назад: предков северных индейцев (ANI), которые имеют большую долю генов, общих с жителями Западной Евразии, и предков южных индейцев (ASI), которые гораздо менее тесно связаны с жителями Западной Евразии (12). Мы использовали древнюю ДНК, чтобы наложить ограничения на генетическую структуру ANI и ASI и, в сочетании с другими данными, сделать выводы о том, когда и где они сформировались. Моделируя современных жителей Южной Азии наряду с древними людьми из мест, находящихся в культурном контакте с IVC, мы вывели вероятную генетическую сигнатуру людей цивилизации долины Инда (IVC), которая достигла своей зрелости на северо-западе Южной Азии в 2600-1900 годах нашей эры. Мы также рассмотрели, когда происходили от степных скотоводов предки (9) смешались с группами населения в Южной Азии и ограничили возможности для того, чтобы определить, какая родословная — степная или иранская — более правдоподобно связана с распространением индоевропейских языков в Южной Азии.
Набор данных и Стратегия анализа
Мы получили полногеномные данные о древней ДНК 523 ранее не изученных древних людей и повысили качество данных о 19 ранее секвенированных людях. Эти люди происходят из трёх обширных географических регионов: 182 человека из Ирана и южной части Центральной Азии, которую мы называем Тураном (современные Туркменистан, Узбекистан, Таджикистан, Афганистан и Кыргызстан), 209 человек из степи и северной лесной зоны, в основном на территории современных Казахстана и России, и 132 человека из северного Пакистана. Древние особи принадлежат к 1) Ирану и Турану эпохи мезолита, меди, бронзы и железа (12000-1 гг. до н.э. из 19 стоянок), включая археологический комплекс Бактрия-Маргиана (BMAC); 2) ранним охотникам-собирателям, использующим керамику, из лесной зоны Западной Сибири, которые, как мы показываем, представляют собой точку в раннем голоцене Северной Евразии и являются ценным источником населения для моделирования родословной жителей Центральной и Южной Азии (6400-3900 гг. до н.э. из 2 стоянок); 3) Скотоводам медного и бронзового веков из центральной степи, в том числе из Казахстана бронзового века (3400-800 гг. до н.э. на 56 стоянках); и 4) самая северная часть Южной Азии, в частности, поздний бронзовый век, железный век и исторические поселения в районах Сват и Читрал на территории современного Пакистана (~1200 г. до н. э. — 1700 г. н. э., 12 мест) (рис. 1, таблица S1, (1, 13)). Мы подготовили образцы в специальных чистых помещениях, извлекли ДНК (14, 15) и создали библиотеки для секвенирования Illumina (16, 17). Мы обогатили библиотеки ДНК, перекрывающими около 1,2 миллиона однонуклеотидных полиморфизмов (SNP) (7, 18, 19), секвенировали продукты на приборах Illumina и провели контроль качества (Таблица S2) (7, 19, 20). Наш окончательный набор данных после объединения с ранее опубликованными данными (7–9, 16, 18, 19, 21–31) охватывает 837 древних особей, прошедших все наши фильтры анализа, которые включали удаление особей, генетически признанных родственниками первой степени других лиц с более высоким охватом (таблица S3), и ограничение 92% особей (таблица S1), которые были представлены по меньшей мере 15 000 целевых SNP, что, как мы обнаружили, было числом, при котором мы начали иметь возможность достоверно оценивать пропорции глубоко расходящихся предков источники. Среднее количество проанализированных SNP на одного человека составило 617 000. Мы также объединили данные полногеномного секвенирования 686 современных людей (Таблица S1) и провели совместный анализ с 1789 современными людьми из 246 этнографически различных групп в Южной Азии, генотипированных по ~600 000 SNP (Таблица S5; (13)) (10, 32, 33).

Мы сгруппировали людей на основе археологической и хронологической информации, используя 269 прямых радиоуглеродных дат, полученных для скелетного материала людей, у которых мы взяли ДНК (Таблица S4). Мы дополнительно сгруппировали людей, которые были генетически неотличимы друг от друга в рамках этих групп, и обозначили отклонения, когда происхождение людей значительно отличалось от других людей, живших в том же месте и в тот же период (13). Для нашего основного анализа мы не включали в него людей, которые были единственными представителями своих родословных, тем самым снижая вероятность того, что наши выводы были сделаны на основе данных отдельных людей с загрязнённой ДНК или неправильно атрибутированным археологическим контекстом. Это также гарантировало, что каждая основная группа анализа была представлена гораздо большим количеством SNP, чем наш минимальный порог в 15 000 на человека. Таким образом, все кластеры, кроме одного, включали по крайней мере одного человека, у которого было более 200 000 однонуклеотидных полиморфизмов, что достаточно для анализа истории популяции с высоким разрешением (19) (исключение составляет пара генетически схожих особей из Гонура, которые не являются предметом основных анализов). Мы используем Курсивом обозначаются генетические группы, а обычным шрифтом — археологические культуры или места.
Чтобы сделать выводы о структуре популяции, мы начали с проведения анализа главных компонент (PCA), проецируя древних людей на паттерны генетической изменчивости современных евразийцев. Эта процедура позволила нам получить значимые ограничения на происхождение даже древних людей с ограниченным охватом, поскольку каждый SNP у каждого человека можно сравнить с большим набором эталонных данных (34–36). Это позволило выявить три основных кластера, тесно связанных с географическими регионами лесной зоны/степи, Ирана/Турана и Южной Азии (рис. 1). Эту закономерность мы воспроизвели в ADMIXTURE при кластеризации без учителя (37). Чтобы проверить, были ли группы древних людей неоднородными по происхождению, мы использовали f4-статистику, чтобы измерить, отличаются ли разные разбиения этих групп на две подгруппы по степени сходства аллелей с третьей группой (используя в качестве базовой группы отдалённо родственную группу). Мы также использовали f3-статистику для проверки на примесь (33). Чтобы смоделировать происхождение каждой группы, мы использовали qpAdm оценивает, соответствует ли тестируемая группа происхождению от заранее заданного количества исходных популяций (по отношению к набору внешних групп), и если да, то оценивает доли предков (7). Сначала мы использовали qpAdm, чтобы попытаться смоделировать группы бронзового века и последующих периодов как смесь семи «дальних» источников, тесно связанных с популяциями бронзового века или отдалённо родственными современными популяциями, для которых у нас есть данные (Вставка 1). В этой статье мы используем термин «земледельцы» для обозначения людей, которые либо выращивали сельскохозяйственные культуры, либо разводили скот, либо и то, и другое. Это определение охватывает не только крупные оседлые общины, но и более мелкие и, вероятно, менее оседлые общины, такие как ранние скотоводы в горах Загрос на западе Ирана, жившие в Гань-Даре. Последние разводили домашних животных, но не выращивали сельскохозяйственные культуры и являются ключевой контрольной группой для этого исследования, поскольку у них был особый генетический профиль, который широко распространился после неолита (9, 24, 38). Мы также определили «проксимальные» модели для каждой группы как смеси предшествующих во времени групп (10). Мы реализовали алгоритм, DATES, для оценки возраста популяционных смешений путем измерения среднего размера сегментов родословной, происходящих из смешивающихся популяций. Этот подход, надежность которого мы проверили с помощью компьютерного моделирования (10), является усовершенствованием по сравнению с методами, не оптимизированными для анализа древней ДНК (33, 39) (надежность этого подхода обусловлена тем, что его молекулярные часы основаны на точно измеренной скорости мейотической рекомбинации у людей (40)). В приложении 2 мы обобщаем результаты этого анализа (мы используем одни и те же заголовки в приложении 2 и в основном тексте, чтобы можно было ссылаться друг на друга), а онлайн-визуализатор данных (1) позволяет интерактивно изучать данные.
Вставка 1. Семь исходных популяций, используемых для моделирования дальнего родства.
| Анатолий_Н | Анатолийские земледельцы: представлены западными анатолийскими земледельцами 7го тысячелетия до н. э. (19) |
| Гандж_дарех_н | Ранние земледельцы Ирана: представлены скотоводами 8го тысячелетия до н. э. из гор Загрос в Иране (9, 24) |
| ВЕХГ | Родственные западноевропейским охотникам-собирателям: представлены западноевропейцами 9го тысячелетия до н. э. (7, 19, 32, 69). (WEHG и EEHG, о которых говорится ниже, в предыдущих исследованиях обозначались как WHG и EHG, но поскольку мы анализируем их вместе с охотниками-собирателями из Азии, мы изменили названия, чтобы указать на европейское происхождение.) |
| ЭЭГ | Родственные восточноевропейским охотникам-собирателям: представлены охотниками-собирателями 6го тысячелетия до н. э. из Восточной Европы (19, 32) |
| WSHG | Родство с западносибирскими охотниками-собирателями: ранее не описанный глубинный источник евразийского происхождения, представленный в этом исследовании тремя особями из лесной зоны Центральной России, датируемыми 6м тысячелетием до н. э. |
| ЭШГ | Восточносибирские охотники-собиратели: представлены охотниками-собирателями 6го тысячелетия до н. э. из региона озера Байкал, чьи предки были тесно связаны с жителями Восточной Азии (26) |
| АХГ | Андаманские охотники-собиратели, родственные современным коренным жителям Андаманских островов (55), которые, по нашему предположению, являются родственниками неандертальцев (древних предков южноиндийцев — AASI) |
Вставка 2. Краткое изложение ключевых выводов.
Иран и Туран
- Градиент уменьшения доли предков, связанных с анатолийскими земледельцами, с запада на восток. В медно-каменном и бронзовом веках в Евразии существовал градиент доли предков с запада на восток — градиент Юго-Западной Азии — с большей долей предков, связанных с анатолийскими земледельцами, на западе и большей долей предков, связанных с WSHG или AASI, на востоке, наложенных на основную долю предков, связанных с ранними иранскими земледельцами. Появление этого градиента по времени совпадает с распространением земледелия в этом регионе, что повышает вероятность того, что люди анатолийского происхождения распространили эту технологию на восток так же, как они помогли распространить её на запад, в Европу.
- Люди, жившие в БМАК, не были основным источником происхождения жителей Южной Азии. Основное население БМАК в значительной степени произошло от предшествующих местных народов эпохи меди, которые, в свою очередь, были тесно связаны с жителями Иранского нагорья и имели мало степного происхождения, распространённого сегодня в Южной Азии.
- Степные скотоводы-кочевники прибыли в Туран к 2100 году до н. э. Мы не обнаружили свидетельств степных скотоводов-кочевников в группах, живших на территории BMAC до 2100 года до н. э., но многочисленные захоронения отдельных людей на этих территориях показывают, что к ~2100–1700 годам до н. э. общины BMAC регулярно взаимодействовали с народами, имеющими такое происхождение.
- Профиль предков, широко распространённый во времена цивилизации долины Инда. Мы задокументировали характерный профиль предков — 45–82% иранских земледельцев и 11–50% AASI (с незначительной примесью анатолийских земледельцев), — который присутствовал в двух местах, находившихся в культурном контакте с культурой долины Инда (IVC). В сочетании с обнаружением того же генетического профиля (в смешанной форме) примерно на тысячу лет позже в долине Сват, существовавшей после ИВК, это свидетельствует о периферийной линии Инда во время расцвета ИВК. Предки этой группы сформировались в результате смешения ~5400–3700 гг. до н. э. В периферийной линии Инда практически нет генетических связей с анатолийскими земледельцами.
Степная и Лесная зона
- Снижение численности населения Северной Евразии после появления земледелия. До распространения земледелия и скотоводства в Северной Евразии существовал градиент с запада на восток, характеризующийся очень разными популяциями охотников-собирателей с возрастающей долей родства с современными жителями Восточной Азии: от западноевропейских охотников-собирателей (WEHG) к восточноевропейским охотникам-собирателям (EEHG), к западносибирским охотникам-собирателям (WSHG) и к восточносибирским охотникам-собирателям (ESHG). Смешение народов вдоль этого генетического градиента и его аналога на юге сформировало пять более поздних клинов после появления земледелия. Три северных из них — это Европейский клин, Кавказский клин и Центральноазиатский клин
- Уникальный генетический профиль, охватывающий территорию от Восточной Европы до Казахстана в бронзовом веке. Мы добавили более ста образцов из ранее описанного генетического кластера Western_Steppe_MLBA, в том числе образцы, связанные с археологическими комплексами шнуровой керамики, срубной культуры, Петровки и Синташты, и характеризующиеся примерно на две трети происхождением от скотоводов ямной культуры (из Кавказской линии) и европейских земледельцев (из Европейской линии). Это позволяет предположить, что данная популяция сформировалась на географической границе этих двух групп в Восточной Европе. Наш анализ показывает, что в центральной степи и Минусинском бассейне в эпоху средней и поздней бронзы западно-степные_палеолитические_люди смешались примерно с 9% ранее проживавших в регионе людей, носивших WSHG, и сформировали особый центрально-степной_палеолитический_кластер, который стал основным каналом распространения в Южную Азию предков скотоводов из ямной культуры.
- Двунаправленная мобильность вдоль горного коридора Внутренней Азии. Начиная с 3-го тысячелетия до н. э. и усиливаясь во 2м тысячелетии до н. э., мы наблюдаем в Центральной степи множество людей, которые жили вдоль горного коридора Внутренней Азии и имели примесь туранской крови, что свидетельствует о движении на север в степи в этот период. К концу 2го тысячелетия до н. э. к этим людям присоединились многочисленные чужеродные группы с восточноазиатскими корнями, которые стали широко распространены в регионе к железному веку (25, 52). Эта родословная также прослеживается в более поздних группах, оказавших известное культурное влияние на Южную Азию, включая гуннов, кушанов и саков, и почти не встречается в двух основных группах предков Южной Азии, что позволяет предположить, что степная родословная, широко распространённая в Южной Азии, произошла от центральноазиатских народов, живших до железного века.
Южная Азия
- Три линии происхождения, сменявшие друг друга во времени в Южной Азии. Мы выделяем уникальное трио исходных популяций, которые соответствуют географически и хронологически разнообразным южноазиатским популяциям бронзового века: смесь AASI, группы Индийской периферии с преимущественно иранским происхождением и Central_Steppe_MLBA. Двусторонние клины, сформировавшиеся в результате смешения этих трёх источников, сменяли друг друга во времени: до 2000 г. до н. э. клины Индо-Арийской периферии не имели степного происхождения, после 2000 г. до н. э. сформировался степной клин, а затем современный индийский клин.
- ASI и ANI возникли в результате смешения жителей периферии Индской культуры с группами, жившими на севере и востоке. Градиент происхождения, частью которого были жители периферии Индской культуры, сыграл ключевую роль в формировании двух основных источников происхождения в Южной Азии: минимум ~55% периферии Индской культуры для ASI и ~70% для ANI. Сегодня в Южной Азии есть группы людей, чьё происхождение очень похоже на статистически реконструированное ASI, что позволяет предположить, что у них есть прямые потомки. Большая часть формирования как ASI, так и ANI произошла во 2м тысячелетии до н. э. Таким образом, события, сформировавшие ASI и ANI, совпали по времени с упадком IV культуры.
- Степное происхождение в Южной Азии в основном связано с мужчинами и непропорционально велико у брахманов. Большая часть степного происхождения в Южной Азии связана с мужчинами, что указывает на асимметричное социальное взаимодействие между потомками степных скотоводов и народами Индийской периферии. Группы, считающие себя традиционно священническими, в том числе традиционные хранители литургических текстов на раннем индоевропейском языке санскрите, как правило (за некоторыми исключениями), имеют более степное происхождение, чем можно было бы ожидать, исходя из смешения ANI-ASI, что является независимым доказательством степного происхождения индоевропейских языков Южной Азии.
Иран и Туран
Линия, идущая с запада на восток, с уменьшающимся количеством предков-земледельцев из Анатолии
Мы изучили генетические изменения, сопровождавшие распространение сельского хозяйства на восток от Ирана, начиная с 7го тысячелетия до н. э. (41, 42). Мы подтвердили предыдущие выводы о том, что у скотоводов, живших в 9м и 8м тысячелетиях до н. э. в горах Загрос на западе Ирана, был характерный для Западной Евразии генетический профиль (9, 27), в то время как более поздние группы, проживавшие в обширном регионе, имели смешанную генетическую принадлежность, включавшую как этот профиль, так и генетическую принадлежность ранних анатолийских земледельцев. Наш анализ показывает, что с запада на восток доля примеси анатолийских земледельцев в медном и бронзовом веках уменьшалась с ~70% в Анатолии до ~31% в восточном Иране и ~7% в дальневосточном Туране (рис. 1; (13), рис. S10, таблица S8–S16). Это говорит о том, что археологически задокументированное распространение общего набора одомашненных растений и животных из разных мест по всему этому региону сопровождалось двусторонним распространением людей и смешением с местными группами, с которыми они сталкивались (3, 41, 43, 44). Мы называем это Юго-Западной азиатской линией. На крайнем востоке Климат Юго-Западной Азии (восточный Иран и Туран) у особей 3-го тысячелетия до нашей эры мы обнаруживаем не только наименьшую долю примеси, связанной с анатолийскими фермерами, но и примесь, связанную с западносибирскими охотниками-собирателями (WSHG) (правдоподобно отражающую примесь от не отобранных групп охотников-собирателей, которые населяли этот регион до распространения в нем иранского происхождения, связанного с фермерами). Это показывает, что происхождение, связанное с Северной Евразией, повлияло на Туран задолго до распространения потомков скотоводов Ямной степи в регионе. Мы можем исключить возможность того, что ямная были источником этой североевразийской родословной, поскольку у них было больше предков, связанных с EEHG-, чем с WSHG-, а также высокая частота гаплогруппы митохондриальной ДНК типа U5a, а также типов гаплогрупп Y-хромосомы R1b или R1a, не представленных в Иране и Туране в этот период ((13), таблица S93-S94).
Люди, жившие на территории BMAC, не были основным источником происхождения южноазиатских народов
В Иране и Туране бронзового века мы получили полногеномные данные для 84 древних людей (3000–1400 гг. до н. э.), которые жили в четырёх городских поселениях Бактрийско-Маргианского археологического комплекса (БМАК) и его непосредственных преемников. Подавляющее большинство этих людей генетически схожи с предыдущими группами в Туране, что согласуется с гипотезой о том, что БМАК сформировался из предшествующих нему городских поселений (5). Мы предполагаем наличие трёх основных генетических источников: происхождение от ранних иранских земледельцев (~60–65 %) и меньшие доли происхождения от анатолийских земледельцев (~20–25 %) и WSHG (~10 %). В отличие от предшествующих им людей бронзового века из Турана, представители кластера BMAC также имели на 2–5 % больше родственных связей (в отдалённом прошлом) с андаманскими охотниками-собирателями (AHG). Это свидетельство распространения генов с севера на юг из Южной Азии согласуется с археологическими данными о культурных контактах между цивилизацией долины Инда и BMAC, а также о существовании торговой колонии IVC на севере Афганистана (хотя у нас нет древней ДНК с этого места) (45), и противоречит нашим qpAdm анализ показывает, что взаимное распространение с севера на юг невозможно обнаружить. В частности, наш анализ показывает, что BMAC и люди, жившие до них в Туране, не могут быть вероятными основными источниками происхождения различных древних и современных народов Южной Азии, поскольку соотношение их анатолийских и иранских фермерских генов слишком велико, чтобы они могли быть вероятным источником происхождения народов Южной Азии (p<0,0001, критерий χ2; (13), рис. S50–S51). Предыдущее исследование (26) соответствовало модели, в которой население Турана медного века использовалось в качестве источника иранских фермерских генов у современных жителей Южной Азии. Таким образом, повышается вероятность того, что люди из BMAC, которые, как правильно предположили авторы, в основном произошли от групп, предшествовавших им в Туране, были основной популяцией-источником для жителей Южной Азии. Однако в этом исследовании были доступны только 2 образца этого периода по сравнению с 36 образцами, о которых мы сообщаем в этом исследовании, и в нём не было древней ДНК людей периода BMAC или каких-либо древних жителей Южной Азии. Благодаря дополнительным образцам мы можем с уверенностью сказать, что ни одна из многочисленных популяций бронзового и медного веков из Турана, для которых у нас есть древняя ДНК, не является источником иранской фермерской родословной в Южной Азии.
Предки степных скотоводов прибыли в Туран к 2100 году до н. э.
Большой размер нашей выборки из Центральной Азии, в том числе из памятников BMAC, является особенно сильной стороной этого исследования. Это позволяет нам выявлять отдельных людей, происхождение которых отличается от происхождения тех, кто жил в то же время и в том же месте, и обнаруживать культурные контакты, которые в противном случае было бы трудно оценить (рис. 2). Примерно в 2300 году до н. э. мы наблюдаем три отклонения в генетических профилях, связанных с BMAC, которые имеют родство с WSHG, и сообщаем о данных, полученных в третьем тысячелетии до н. э. в трёх местах в Казахстане и одном в Кыргызстане, которые могут быть их источниками (родственное происхождение было обнаружено у представителей ботайской культуры, живших примерно в 3500 году до н. э. (26)). Ямная родословная появилась к 2100 году до н. э., поскольку с 2100 по 1700 год до н. э. мы наблюдаем отклонения от нормы в трёх местах, связанных с BMAC, у которых родословная в конечном счёте происходит от скотоводов Западной_Степи_EMBA в характерной смешанной форме, типичной для многих степных групп среднего и позднего бронзового века (примерно две трети родословной имеют Западно-Степное_EMBA происхождение, а остальная часть соответствует происхождению от европейских земледельцев). Таким образом, наши данные документируют движение предков на юг, в конечном счете произошедших от скотоводов Ямной степи, которые распространились в Центральную Азию на рубеже 2-готысячелетия до нашей эры.

Профиль предков , широко распространенный во времена цивилизации долины Инда
Мы документируем 11 выбросов — 3 с радиоуглеродными датами между 2500-2000 годами до н.э. из стоянки BMAC в Гонуре и 8 с радиоуглеродными датами или археологическими контекстными датами между 3300 г. до н.э. и 2000 г. до н.э. из восточноиранской стоянки Шахри-и-Сохта — в которых наблюдалась повышенная доля предков, связанных с AHG (диапазон 11-50%), а остальные – характерная смесь предков, связанных с иранскими фермерами и WSHG (~ 50-89%).). В отличие от основных кластеров BMAC (~20–25% родства с анатолийскими фермерами) и Шахр-и-Сохта (~16–21%), эти выбросы не имели заметного родства с анатолийскими фермерами, что позволяет нам отвергнуть как BMAC, так и основной кластер Шахр-и-Сохта в качестве их источников (p<10−7, критерий χ2; (13), Таблица S83). Без древней ДНК людей, похороненных в культурных контекстах ИВК, мы не можем с уверенностью утверждать, что генетический градиент, представленный этими 11 исключительными особями, который мы называем Индийской периферийной линией, также был характерен для ИВК. Тем не менее, наш результат дает шесть косвенных доказательств в пользу этой гипотезы. (i) У этих особей не было обнаруживаемой родословной, связанной с анатолийскими фермерами, что позволяет предположить, что они происходят из групп, расположенных дальше на восток вдоль линии Анатолия-Иран с уменьшающейся родословной, связанной с анатолийскими фермерами, чем у любых особей, отобранных нами за этот период. (ii) Все 11 выбросов имели повышенную долю предков, связанных с AHG-, а двое несли гаплогруппу H1a1d2 Y-хромосомы, которая сегодня в основном встречается в южной Индии. (iii) И в Гонуре, и в Шахр-и-Сохте есть археологические свидетельства обмена с IVC (46, 47), и все индивиды, которых мы напрямую датировали, относятся ко времени зрелой IVC. (iv) Несколько индивидов из Шахр-и-Сохты были похоронены с артефактами, стилистически связанными с Белуджистаном в Южной Азии, в то время как захоронения, связанные с другими предками, не имели таких связей (13). (v) В нашем моделировании 11 выбросов подходят в качестве основного источника происхождения 86 древних особей из культур после IVC, живущих вблизи истоков реки Инд ~ 1200-800 гг. до н.э., а также разнообразных современных жителей Южной Азии, тогда как никакие другие древние генетические кластеры из Турана не подходят в качестве источников для всех этих групп ((13), рис. S50). (vi) Предполагаемая дата смешения иранских фермеров и AHG- предков в выбросах на несколько тысячелетий раньше того времени, когда они были обнаружены. жил (71 ± 15 поколений, что соответствует 95% доверительному интервалу ~ 5400-3700 гг. до н.э. при условии, что 28 лет на поколение (13, 48). Таким образом, AHG и связанные с иранскими земледельцами группы контактировали задолго до появления зрелой ИВК в ~2600–1900 гг. до н. э., как и следовало ожидать, если бы генетический градиент был характерной чертой группы, жившей в долине Инда во времена ИВК.
Степная и Лесная зона
Резкие изменения в происхождении в Евразии установились после появления земледелия
Поздние охотники-собиратели из Северной Евразии располагаются вдоль градиента охотников-собирателей с запада на восток, по мере увеличения родства с жителями Восточной Азии (рис. 3). В эпоху неолита и меди охотники-собиратели, жившие в разных точках вдоль этого клина, смешивались с людьми, чьи предки жили в разных точках вдоль южного клина, образуя пять более поздних клинов, два из которых находились на юге (клины Юго-Западной Азии и Индийской периферии, которые описаны в предыдущем разделе), а три — в Северной Евразии (рис. 3). Дальше всего на запад в степной и лесной зонах простиралась европейская клинальная популяция, сформировавшаяся в результате распространения земледельцев из Анатолии после ~7000 г. до н. э. и смешения с западноевропейскими охотниками-собирателями (19). В дальневосточной Европе на широтах, охватывающих Чёрное и Каспийское моря, простиралась кавказская клинальная популяция, состоящая из смеси восточноевропейских охотников-собирателей и иранских земледельцев с дополнительным анатолийским земледельческим происхождением в некоторых группах (49). К востоку от Урала мы обнаруживаем Центральноазиатскую клину с представителями WSHG на одном конце и представителями медного и раннего бронзового веков из Турана на другом.

Уникальный генетический профиль, распространённый от Восточной Европы до Казахстана в бронзовом веке
Примерно с 3000 г. до н. э. генетические профили многих групп населения Евразии претерпели изменения в результате распространения ямной степной палеолитической культуры (Western_Steppe_EMBA) из её истоков в Кавказской складке (9, 49) на обширную территорию, простирающуюся от Венгрии на западе до гор Алтая на востоке (7, 8) (рис. 3). В течение следующих двух тысячелетий эта ветвь распространялась дальше, смешиваясь с местными группами, и в конечном итоге достигла атлантических берегов Европы на западе и Южной Азии на юго-востоке. Источником западно-степного_EMBA происхождения, которое в конечном итоге достигло Центральной и Южной Азии, была не первоначальная экспансия на восток, а вторичная экспансия, в которой участвовала группа, у которой было ~67% западно-степного_EMBA происхождения и ~33% происхождения из точки на европейской линии (8) (рис. 3). Мы подтверждаем предыдущие выводы о том, что в эту группу входили представители археологических комплексов шнуровой керамики, срубной культуры, петровской культуры и синташтинской культуры, которые проживали на обширной территории от границы Восточной Европы до северо-западного Казахстана (8, 19, 21), и наш набор данных включает более сотни представителей этого кластера Western_Steppe_MLBA. Мы также обнаруживаем ещё один кластер, Central_Steppe_MLBA, который отличается от Western_Steppe_MLBA (p=7×10−6 по qpAdm) наличием ~9% дополнительной родословной, полученной от скотоводов бронзового века из центральной степи, в основном имеющих родословную, связанную с WSHG (Central_Steppe_EMBA). Таким образом, люди с родословной Western_Steppe_MLBA смешивались с местным населением по мере продвижения на восток и юг.
Двунаправленная мобильность По Горному коридору Внутренней Азии
Как и в Иране/Туране, отдельные индивидуумы предоставляют важную информацию о взаимодействии людей.
Во-первых, наш анализ 50 человек с кладбища синташтинской культуры в Каменном Амбаре V выявил несколько групп «выбросов», которые, согласно прямым радиоуглеродным датировкам, были современниками основной группы, но генетически отличались от неё, что указывает на космополитичность этого места (рис. 2). У одной группы аутсайдеров была повышенная доля центрально-степного_EMBA (в основном WSHG-) происхождения, у другой — повышенная доля западно-степного_EMBA (ямной) происхождения, а у третьей — повышенная доля EEHG- происхождения.
Во-вторых, в центральной степи (современный Казахстан) у индивида, жившего в 2800–2500 гг. до н. э. на одном из участков, и у индивидов, живших в 1600–1500 гг. до н. э. на трёх участках, наблюдается значительная примесь генов иранских земледельцев, что хорошо согласуется с основным кластером BMAC, демонстрируя миграцию генов с севера Турана в степь в то же время, когда на юг через Туран в Южную Азию мигрировали представители Central_Steppe_MLBA-. Таким образом, археологически задокументированное распространение материальной культуры и технологий как на север, так и на юг вдоль Внутреннеазиатского горного коридора (50, 51), которое началось ещё в середине 3го тысячелетия до н. э., было связано с масштабными перемещениями людей (рис. 2).
В-третьих, мы наблюдаем особей из степных стоянок (Красноярск), датируемых ~ 1700-1500 годами до н.э., которые на ~ 25% происходят из источника, связанного с восточноазиатами (хорошо смоделированного как ESHG), а остальная часть лучше всего смоделирована как Western_Steppe_MLBA. К позднему бронзовому веку примесь, связанная с ESHG, стала повсеместной, что подтверждается нашим временным срезом из Казахстана, а также данными древней ДНК железного века и более поздних периодов в Туране и Центральной степи, включая скифов, сарматов, кушанов и гуннов (25, 52). Таким образом, эти археологические культуры, существовавшие с 1го тысячелетия до н. э. по 1е тысячелетие н. э. и оказавшие заметное культурное и политическое влияние на Южную Азию, не могут быть важными источниками степной кочевой родословной, распространённой в Южной Азии в наши дни (поскольку у современных жителей Южной Азии слишком мало восточноазиатской родословной, чтобы они могли происходить от этих групп). Это пример того, как генетические данные могут опровергнуть сценарии, которые кажутся правдоподобными только на основании археологических и исторических свидетельств ((13), рис. S52). Вместо этого наш анализ показывает, что единственным вероятным источником степной родословной являются степные группы среднего и позднего бронзового века, которые не только подходят на роль источника для Южной Азии, но и, как мы выяснили, распространились в Туран и смешались с представителями BMAC в Казахстане в этот период. В совокупности эти результаты указывают на узкий временной промежуток (первая половина второго тысячелетия до н. э.), когда степная родословная, широко распространённая сегодня в Южной Азии, должна была появиться.
Геномное формирование человеческих популяций в Южной Азии
Три родословные линии, которые сменяли друг друга во времени в Южной Азии
Предыдущие исследования показали, что южноазиатские народы имеют общие корни с древними группами, проживавшими на севере Евразии и в Иране, в Восточной Азии и в Австралазии (9). Здесь мы описываем процесс, в ходе которого эти древние группы смешались и образовали более поздние группы.
Мы начнём с периода до 2000 г. до н. э., периферийной зоны долины Инда, описанной в предыдущем разделе и обнаруженной в 11 выбросах из двух мест, находившихся в культурном контакте с цивилизацией долины Инда (рис. 4). Мы можем смоделировать всех людей, входящих в эту клину, как смесь двух исходных популяций: один конец клины соответствует полностью AHG-связанной популяции, а другой соответствует 90% иранских земледельцев и 10% WSHG-связанной популяции (рис. 4, (13)). Люди, входящие в периферийную клину Инда, составляют большинство предков современных жителей Южной Азии. С помощью формального моделирования мы демонстрируем, что именно вклад представителей Индской периферийной культуры в генофонд более поздних жителей Южной Азии, а не поток генов с запада, принесший на Иранское плато уникальную для Южной Азии родословную, объясняет высокую степень родства между современными жителями Южной Азии и иранцами раннего голоцена (9, 13, 27).

Затем мы охарактеризовали степной климат 2000 г. до н.э., представленный в нашем анализе 117 особями, датируемыми 1400-1700 гг. до н.э. из районов Сват и Читрал на самом севере Южной Азии (рис. 2, рис. 4). Мы обнаружили, что можем совместно смоделировать всех особей в Степной группе как смесь двух источников, хотя и отличающихся от двух источников в более ранней группе. Один конец соответствует точке вдоль Индийской периферической линии. Другой конец соответствует смеси примерно 41% Центрально-степного происхождения и 59% от подгруппы Индийской периферической линии с относительно высоким уровнем иранского происхождения ((13), рис. S50).
Чтобы понять, как сформировалась современная индийская линия, мы искали тройки популяций, которые могли бы служить источниками для различных современных южноазиатских групп, а также народов степной линии. Все подходящие модели включают в качестве источников Central_Steppe_MLBA (или группу со схожим профилем происхождения), группу Indus Periphery Cline и либо AHG, либо подгруппу Indus Periphery Cline с относительно высоким уровнем AHG-родственных связей (13), рис. S51). Совместный анализ 140 различных южноазиатских групп (10), которые распределяются по градиенту в методе главных компонент (13), показывает, что, несмотря на наличие трёх основных источников, как и в случае с двумя предыдущими клинами, подавляющее большинство групп на современном индийском клине можно смоделировать как смесь двух популяций, которые произошли от трёх предыдущих. Хотя у нас нет данных о древней ДНК ни одной из двух статистически реконструированных исходных популяций для современной индейской линии, ASI или ANI, в дальнейшем мы проанализируем наши данные о древней ДНК в сочетании с современными данными, чтобы определить точное происхождение ASI и ограничить распространение ANI.
ASI и ANI возникли в результате смешения жителей периферии долины Инда с группами, жившими на севере и востоке
Чтобы получить представление о формировании ASI, мы экстраполировали до наименьшей теоретической крайности, связанной с Западной Евразией, современную индийскую линию, установив в нашей модели нулевую долю степного_народа_MLBA. Мы оцениваем, что минимум 55% предков происходят от людей, живших на периферии Инда (представляя периферию Инда человеком, у которого больше всего предков, связанных с иранскими земледельцами, которого мы называем Indus_Periphery_West), а остальная часть предков происходит от группы, связанной с AHG (13). Мы обнаружили, что несколько племенных групп на юге Индии имеют ~0% предков из Центральной степи (13). Тот факт, что эти люди соответствуют наиболее экстремальной из возможных позиций для ASI, не только показывает, что почти прямые потомки ASI сегодня живут в Южной Азии, но и позволяет нам сделать точное заявление о происхождении ASI. В частности, тот факт, что у них есть значительная доля иранской фермерской крови (через Индийскую периферию), опровергает более ранние предположения о том, что ASI не имеют родственных связей с западноевразийцами (11). Использование DATES мы оцениваем в среднем 107 ± 11 поколений с момента смешения иранских земледельческих и AHG-родственных групп в одной из этих групп: Паллияр. Это соответствует 95-процентному доверительному интервалу 1700–400 гг. до н. э. при условии, что на одно поколение приходится 28 лет (53). Таким образом, ASI не были полностью сформированы во времена ИВК и, должно быть, продолжали формироваться путём смешения после её упадка, когда материальная культура, характерная для ИВК, распространилась на восток (54), а индская периферийная группа смешалась с людьми, менее родственными западным евразийцам.
Мы также получили дополнительные доказательства позднего (бронзового века) формирования ASI, построив график смешения с помощью qpGraph и смоделировав Palliyar и Juang (австроазиатскую группу в Индии с низким уровнем родства с западноевразийцами) (рис. 5). График соответствует компоненту южноазиатского происхождения без родственных связей с Западной Евразией (AASI — «древние предки из Южной Азии») как азиатской линии, отделившейся примерно в то же время, когда предки из Восточной Азии, с Андаманских островов и австралийские аборигены отделились друг от друга, что согласуется с гипотезой о том, что восточноазиатские и южноазиатские линии произошли от распространения на восток, которое за короткий промежуток времени привело к появлению линий, ведущих к AASI, жителям Восточной Азии, охотникам-собирателям с Андаманских островов и австралийцам (55) (рис. 5). Джуанг не может быть сопоставлен со смесью родословной азиатов и родословной, связанной с носителями австроазиатского языка, и вместо этого может быть сопоставлен только путем моделирования дополнительной родословной из ААСИ, показывая, что в то время, когда австроазиатские группы формировались в Южной Азии, также присутствовали группы с меньшим происхождением, связанным с иранским сельским хозяйством, чем в АСИ. Предполагается , что австроазиатские языки распространились в Южной Азии в 3 веке н. э.Третье тысячелетие до н. э. (на основе систем земледелия на холмах, которые, как предполагается, связаны с распространением австроазиатских языков (42), и, таким образом, генетический профиль юангов является независимым доказательством позднего (бронзового века и, вероятно, после IV века до н. э.) формирования австроазиатской языковой семьи.

Чтобы пролить свет на формирование статистически реконструированного ANI, мы возвращаемся к временному разрезу долины Сват, который сформировал Степной склон после 2000 года до нашей эры. Современный индийский климат пересекает Степной климат в месте, близком к положению калашей, группы на северо-западе Южной Азии с самой высокой долей предков ани (56) (рис. 4). Оценка примеси в калашах, основанная на ДАТАХ, составляет 110 ± 12 поколений (56), что предполагает, что дата образования ANI после IVC совпадает с датой образования ASI после IVC. Дальнейшие доказательства интеграции степных предков в Южную Азию после IVC поступают от древних особей на Степном склоне (вдоль которого теоретически могли сформироваться ани), чья примесь степных предков также относится к периоду после IVC. В частности, мы оцениваем, что примесь в популяциях позднего бронзового и железного веков в районе Сват на севере Южной Азии произошла в среднем за 26 поколений до того, как они жили, что соответствует 95-процентному доверительному интервалу ~1900–1500 гг. до н. э. Эта временная шкала появления степных предков в регионе согласуется с нашими наблюдениями за 6 отдельными особями в Туране, которые жили в период с ~2000 по 1500 год до н. э. и имеют эту родословную в смешанной форме (рис. 2), а также с нашим открытием, что Y-хромосома R1a, связанная с родословной Центральной_Степи_MLBA в Южной Азии, также присутствует у жителей района Сват в эпоху поздней бронзы и железного века (2 копии).
В совокупности эти результаты показывают, что ни одна из двух основных популяций, послуживших источником современного индийского генома, ANI и ASI, не была полностью сформирована до начала 2го тысячелетия до н. э.
Степное происхождение в Южной Азии в основном связано с мужчинами и непропорционально велико у брахманов
У особей позднего бронзового и железного веков долины Сват мы обнаруживаем значительно более низкую долю степной примеси в Y-хромосоме (только 5% из 44 Y-хромосом подтипа R1a-Z93, которая встречается со 100% частотой у Central_Steppe_MLBA самцов) по сравнению с 20% в аутосомах (Z = -3,9 для дефицита у самцов при упрощающем предположении, что все Y-хромосомы не связаны друг с другом с момента смешения и, следовательно, статистически независимы), документируя, как степное происхождение было включено в аутосомы. в эти группы попали в основном женщины (рис. 4). Однако в разных частях Южной Азии гендерная предвзятость была разной, поскольку у современных жителей Южной Азии мы наблюдаем обратную картину: избыток Y-хромосомной ДНК, связанной с Центральной степью по сравнению с аутосомами (Z = 2,7 для избытка у мужчин) (13, 57) (рис. 4). Таким образом, распространение степных пастушеских линий среди предков современных жителей Южной Азии происходило в основном через мужчин. Это смещение в сторону более высокого уровня похоже на то, что было зафиксировано при проникновении степных народов в Иберию на крайнем западе Европы, хотя смещение в этом случае менее выражено (58).
Наш анализ степной родословной также выявил 6 групп с очень значительным превышением ожидаемого соотношения Central_Steppe_MLBA- и Indus_Periphery_West- родословных по сравнению с моделью на уровне Z < −4,5. Два самых сильных сигнала были у Brahmin_Tiwari (Z = −7,9) и Bhumihar_Bihar (Z = −7,0). В целом наблюдается заметное обогащение в группах, которые считают себя традиционно жреческими: 5 из 6 групп с Z < −4,5 были браминами или бхумихарами, хотя они составляют всего 7–11% из 140 проанализированных групп (p<10−12 по критерию χ2 при условии, что все группы эволюционировали независимо). Мы предупреждаем, что это не формальный тест, поскольку существует неизвестная степень общего происхождения между группами, поскольку они сформировались в результате смешения, а также потому, что наши решения о том, какие группы включать в анализ, не были приняты вслепую. Например, мы исключили четыре группы «католических браминов» с убедительными доказательствами существенного общего происхождения в прошлом тысячелетии (10), что делает их статистически независимыми (Таблица S5, Рис. 4 (13)). Тем не менее, тот факт, что у традиционных хранителей литургии на санскрите (брахманов), как правило, больше степного происхождения, чем можно было бы предположить, исходя из простой модели смешения ASI-ANI, является независимым доказательством, помимо характерного профиля происхождения, общего для Южной Азии и Восточной Европы бронзового века, отражающего общие черты балто-славянских и индоиранских языков (59), степного происхождения индоевропейских языков Южной Азии до ~2000 г. до н. э.
Обсуждение
Наш анализ показывает, что в голоцене в регионе Южной Азии существовало по меньшей мере три генетических градиента. До ~2000 г. до н. э. существовала периферийная зона Инда, состоящая из людей с разной долей иранской фермерской и AASI-родственной ДНК, что, по нашему предположению, было характерной чертой многих представителей IVC. АЗИ сформировались после 2000 г. до н.э. как смесь части этой линии с выходцами из Южной Азии с более высокой долей родства, связанного с ААСИ. Примерно между 2000 и 1000 годами до н.э. люди, имевшие в основном родословную Central_Steppe_MLBA, распространились в сторону Южной Азии, смешавшись с людьми вдоль периферии Инда, образовав Степной регион. Многочисленные точки вдоль степного клина представлены особями из временного ряда долины Сват, и статистически мы обнаруживаем, что АНИ, одна из двух основных популяций-источников в Южной Азии, может соответствовать степному клину. После 2000 г. до н. э. смешение популяций АСИ и АНИ привело к формированию современного индийского клина, который сегодня представлен различными группами в Южной Азии (рис. 4).
Наше открытие, основанное на неравновесии сцепленных гаплогрупп ((13), рис. S59), что смешение, сформировавшее Индийскую периферийную клину, произошло примерно в 5400–3700 гг. до н. э. — по крайней мере, за тысячу лет до формирования зрелой IVC, — указывает на две возможности. Одна из них заключается в том, что происхождение иранских земледельцев в этой группе было характерно для охотников-собирателей долины Инда так же, как и для охотников-собирателей Северного Кавказа и Иранского нагорья. Наличие такой родословной у охотников-собирателей из пещер Белт и Хоту на северо-востоке Ирана повышает вероятность того, что такая родословная могла быть у охотников-собирателей, живших дальше на восток. Альтернативное предположение состоит в том, что такая родословная отражает переселение в Южную Азию с Иранского нагорья людей, сопровождавшее распространение на восток сельского хозяйства, основанного на выращивании пшеницы и ячменя, а также разведении коз и овец, ещё в 7м тысячелетии до н. э. и формирование ранних фермерских поселений, таких как Мергарх на холмах, окружающих долину Инда (60, 61). Однако это противоречит наблюдению о том, что У жителей Индо-Арийской периферии было мало общего с анатолийскими земледельцами, что тесно связано с распространением земледелия на восток в нашем наборе данных. Таким образом, хотя наш анализ подтверждает идею о том, что распространение на восток анатолийской фермерской генетической линии было связано с распространением сельского хозяйства на Иранском нагорье и в Туране, наши результаты не подтверждают крупномасштабные перемещения населения с Ближнего Востока в Южную Азию после ~6000 г. до н. э. (времени, после которого все древние люди из Ирана в наших данных имеют анатолийскую фермерскую генетическую линию, хотя у жителей Южной Азии её очень мало). Языки в догосударственных обществах обычно распространялись в результате переселения людей (62), и поэтому отсутствие значительного количества анатолийских предков-земледельцев в Индийской периферийной зоне позволяет предположить, что индоевропейские языки, на которых сегодня говорят в Южной Азии, вряд ли возникли в результате распространения земледелия из Западной Азии.
Наши результаты не только опровергают теорию о том, что индоевропейские языки Южной Азии произошли от языков Иранского нагорья, но и подтверждают теорию о том, что эти языки распространились из степей. В то время как древняя ДНК свидетельствует о движении степных скотоводов на запад, что, вероятно, способствовало распространению многих индоевропейских языков в Европе (7, 8), цепочка передачи этих языков в Южную Азию неясна из-за отсутствия соответствующей древней ДНК. Наше наблюдение за распространением степного_народа_эпохи_бронзы в Южной Азии в первой половине 2го тысячелетия до н. э. подтверждает это и особенно примечательно тем, что даёт правдоподобное генетическое объяснение языковому сходству между балто-славянской и индо-иранской подгруппами индоевропейских языков, которые, несмотря на большое географическое расстояние между ними, имеют общие инновации сатемской культуры и законы рунического письма (63). Если переселение людей из степей в этот период было каналом распространения индоевропейских языков в Южной Азии, то поразительно, что в материальную культуру центральной части степей и Южной Азии в эпоху средней и поздней бронзы (то есть после середины II тысячелетия до н. э.) было внесено так мало сходства. Действительно, различия в материальной культуре настолько существенны, что некоторые археологи не видят никаких доказательств связи. Однако отсутствие связей с материальной культурой не является доказательством против распространения генов, как было продемонстрировано на примере культуры колоколовидных кубков, которая зародилась в основном в Западной Европе, но в Центральной Европе была связана со скелетами, которые на ~50% состояли из генов, родственных скотоводам ямной культуры (18). Таким образом, в Европе мы имеем однозначный пример того, как люди степного происхождения оказывали глубокое демографическое влияние на регионы, в которые они проникали, перенимая важные аспекты местной материальной культуры. Наши результаты свидетельствуют о подобном явлении в Южной Азии, где у местного населения, подвергшегося аккультурации, встречается до ~20% Western_Steppe_EMBA-в соответствии с нашим моделированием (через группы Central_Steppe_MLBA) (рис. 3). Наш анализ также предоставляет второе доказательство связи между степным происхождением и индоевропейскими языками. Степное происхождение в группах, которые считают себя традиционно жреческими, поражает, поскольку некоторые из этих групп, в том числе брахманы, являются традиционными хранителями литературы, написанной на раннем санскрите. Возможное объяснение состоит в том, что приток представителей степной_группы_MLBA в Южную Азию в середине II тысячелетия до н. э. создал метапопуляцию с разной долей степного происхождения, при этом люди с большим степным происхождением (или с меньшим смешением с группами периферийной линии Инда) были более тесно связаны с индоевропейской культурой. Из-за сильной эндогамии, которая на протяжении тысячелетий изолировала группы от соседей (7), некоторые из этих популяций сохранились в Южной Азии среди современных носителей индоевропейских текстов.
Наши результаты также проливают свет на происхождение второй по величине языковой группы в Южной Азии — дравидийской. Сильная корреляция между происхождением ASI и современными дравидийскими языками позволяет предположить, что ASI, которые, как мы показали, сформировались как группы с происхождением, типичным для периферийной зоны Инда, мигрировали на юг и восток после упадка IVC и смешались с группами с более высоким происхождением AASI, которые, скорее всего, говорили на раннем дравидийском языке. Возможный сценарий, объединяющий генетические данные с археологическими и лингвистическими, заключается в том, что протодравидийские языки распространились среди народов ИВК вместе с компонентом индо-арийской периферии ASI. Негенетические доказательства того, что дравидийские языки произошли от ИВК, включают современное географическое распространение этих языков (на юге Индии и юго-западе Пакистана), а также предположение, что некоторые символы на древних печатях из долины Инда обозначают дравидийские слова или названия (64, 65). Альтернативная версия заключается в том, что протодравидийский язык распространился примерно на половине территории древних индийцев, которая не относится к Индийской периферии, а находится на юге и востоке (на полуострове Южная Азия). Южный сценарий согласуется с реконструкцией протодравидийских терминов, обозначающих флору и фауну, уникальные для полуострова Индостан (66, 67).
В заключение мы отмечаем примечательную параллель между предысторией Южной Азии и Европы. В обоих регионах происходили обмены между людьми, родственными жителям Юго-Западной Азии, и местными жителями; смешение этих групп привело к появлению Индийской периферии в Южной Азии и Европейской периферии в Европе. На обоих субконтинентах люди, прибывшие в 3м и 2м тысячелетиях до н. э. и произошедшие от смешения народов, родственных скотоводам ямной культуры и европейским земледельцам, смешивались с местным населением: в Южной Азии, образуя АНИ, а в Европе — группы, подобные культурам колоколовидных кубков. В обоих случаях смешение этих смешанных популяций — как с примесью степных скотоводов, так и без неё — определяет современные генетические линии в обоих регионах (рис. 3). Однако существуют также значительные различия между распространением предков в бронзовом веке и неолите на двух субконтинентах. Во-первых, максимальная доля местных предков выше в Южной Азии (AASI — до ~60%), чем в Европе (WEHG — до ~30%) (7), что может отражать более сильные экологические или культурные барьеры для расселения людей в Южной Азии, чем в Европе, что давало ранее сформировавшимся группам больше времени для адаптации и смешения с прибывающими группами. Второе отличие заключается в меньшей доле степных скотоводов в Южной Азии, чем в Европе, их более позднем прибытии примерно на 500–1000 лет и меньшем преобладании мужского пола в смешении. Эти факторы помогают объяснить сохранение значительной доли неиндоевропейских языков среди населения современной Южной Азии. Ситуация в Южной Азии чем-то напоминает ситуацию в Средиземноморской Европе, где доля степного происхождения значительно ниже, чем в Северной и Центральной Европе (рис. 3), и где в классические времена были распространены многие неиндоевропейские языки (68). Дальнейшие исследования древней ДНК из Южной Азии и лингвистически родственного ей иранского мира расширят и дополнят представленную здесь модель.
Материалы и методы
Лабораторная работа по древней ДНК.
Для элементов скелета, которые мы не смогли транспортировать с мест раскопок, мы просверлили отверстия непосредственно в костях, в основном в частях каменистой кости, отвечающих за внутреннее ухо, используя метод отбора образцов из основания черепа (CBD) (70). Подавляющее большинство элементов скелета были подготовлены в специальных чистых помещениях для работы с древней ДНК в Гарвардской медицинской школе, Университетском колледже Дублина, Венском университете или Институте эволюционной антропологии Общества Макса Планка в Лейпциге, Германия, путём сверления или пескоструйной обработки для отделения фрагмента кости с последующим фрезерованием (Таблица S1, Таблица S2).
Все молекулярные исследования, кроме исследования одного индивида (Дарра-и-Кур), проводились в Гарвардской медицинской школе (HMS). В HMS мы извлекали ДНК с помощью метода, оптимизированного для сохранения небольших фрагментов ДНК. Мы применяли этот метод как вручную с использованием силикатных центрифужных колонок (565 библиотек) (14, 15), так и с помощью роботизированных манипуляторов с использованием магнитных шариков с силикатным покрытием и буфера D (149 библиотек) (71). Мы преобразовали ДНК в форму, пригодную для секвенирования, с помощью протокола подготовки двухцепочечных библиотек (711 библиотек) и протокола подготовки одноцепочечных библиотек (3 библиотеки) (72). Для всех двухцепочечных библиотек, кроме четырёх, мы предварительно обработали их смесью ферментов урицил-ДНК-гликозилазы (UDG) и эндонуклеазы VIII (USER, New England Biolabs), чтобы уменьшить количество характерных ошибок при замене цитозина на тимин в древней ДНК, за исключением концевого основания (17). Остальные четыре библиотеки не подвергались предварительной обработке USER (73). Три одноцепочечные библиотеки также подвергались предварительной обработке USER таким образом, что это приводило к аналогичному повреждению в виде неэффективного удаления урацила в концевой базе (72). Мы подготовили большинство двухцепочечных библиотек (n=524) с помощью роботизированного манипулятора для работы с жидкостями, заменив колонки MinElute, которые использовались для очистки реакций при ручной обработке, магнитными шариками с покрытием из диоксида кремния при роботизированной обработке, а очистку ПЦР с помощью колонок MinElute в конце подготовки библиотеки — шариками SPRI (74, 75). Мы обогатили все библиотеки как последовательностями, перекрывающими митохондриальную ДНК (76), так и последовательностями, перекрывающими около 1,24 миллиона ядерных мишеней. Мы провели два раунда обогащения для этих целей (7, 19, 20) либо в двух независимых экспериментах по захвату, либо вместе. После индексирования продуктов обогащения таким образом, чтобы каждой библиотеке был присвоен уникальный индекс и комбинация (77), мы секвенировали обогащённые продукты на приборе Illumina NextSeq500 с использованием наборов v.2 150 циклов для 2×76 циклов и 2×7 циклов (2×8 для одноцепочечных библиотек) и секвенировали до тех пор, пока ожидаемое количество дополнительных SNP на 100 дополнительных пар прочитанных фрагментов не стало меньше 1. Мы также секвенировали методом дробовика двухцепочечные библиотеки, чтобы оценить долю последовательностей, которые соответствуют геному человека.
Чтобы проанализировать данные, мы начали с сортировки пар считываний путём поиска ожидаемых идентификационных индексов и штрихкодов для каждой библиотеки, допуская в каждом случае одно отклонение от ожидаемой последовательности. Мы удалили адаптеры и объединили последовательности, требующие перекрытия в 15 пар оснований (допуская одно отклонение), взяв за основу наиболее качественное основание в объединённом сегменте. Мы сопоставили полученные последовательности с эталонной последовательностью hg19 человека (GRCh37, версия, используемая в проекте «1000 геномов» (78)), используя команду samse в BWA (79) (версия 0.6.1). Мы удалили повторяющиеся последовательности (относящиеся к одной и той же позиции в геноме и имеющие одну и ту же пару штрихкодов) и объединили библиотеки, относящиеся к одному и тому же образцу (объединяя образцы, если генетические данные указывали на то, что несколько образцов были получены от одного и того же индивида). Для каждого индивида мы ограничились последовательностями, прошедшими фильтры (не перекрывающими известные полиморфизмы со вставками/вырезами и имеющими минимальное качество картирования 10), и удалили два нуклеотида с конца каждой последовательности, чтобы уменьшить количество артефактов дезаминирования. Мы также ограничились данными о последовательностях с минимальным качеством оснований 20. Чтобы представить каждого человека в каждой позиции SNP, мы случайным образом выбрали одну последовательность (если она была доступна).
Для Дарра-и-Кур мы проанализировали библиотеку одноцепочечной ДНК (L5082) в Институте эволюционной антропологии Общества Макса Планка (MPI-EVA) в Лейпциге, Германия, созданную в рамках предыдущего исследования (80). В предыдущем исследовании анализировалась только митохондриальная ДНК, а для текущего исследования мы обогатили библиотеку последовательностями, перекрывающими ту же панель из примерно 1,2 миллиона ядерных мишеней, с помощью двух раундов гибридизационного захвата (7, 19, 20). Мы секвенировали обогащённые библиотеки на 2 дорожках платформы Illumina HiSeq2500 в конфигурации с двойным индексом (2×76 циклов) (77) и определяли аллели с помощью FreeIbis (81). Мы объединяли перекрывающиеся парные концы и обрезали их с помощью leeHom (82). Мы использовали BWA для выравнивания последовательностей с эталонным геномом человека hg19 (GRCh37) (79). Мы сохраняли последовательности, которые идеально соответствовали ожидаемой комбинации индексов, для последующего анализа.
Мы оценивали подлинность древней ДНК, измеряя степень повреждения первого нуклеотида. Мы отмечали людей как потенциально загрязнённых, если у них было менее 3% замен цитозина на тимин в первом нуклеотиде для библиотеки, обработанной UDG, и менее 10% замен для библиотеки, не обработанной UDG. Мы использовали contamMix для проверки на загрязнение на основе полиморфизма митохондриальной ДНК (83) и ANGSD для проверки на загрязнение на основе полиморфизма Х-хромосомы у мужчин (84).
Радиоуглеродный анализ.
Мы получили 269 радиоуглеродных (14C) дат для костей с помощью масс-спектрометрии с ускорением (AMS) (Таблица S3). Большинство из них (n=242) были получены в радиоуглеродной лаборатории Университета штата Пенсильвания (PSU), и здесь мы приводим описание методологии подготовки образцов в PSU (методы, используемые в других лабораториях, общедоступны, и мы отсылаем читателей к литературе по этим методикам). Возможные загрязняющие вещества (консерванты и адгезивы) удаляли обработкой ультразвуком всех образцов костей при последовательных промывках метанолом, ацетоном и дихлорметаном марки ACS в течение 30 минут при комнатной температуре с последующей трехкратной промывкой в нанопорошковой воде для полоскания. Костный коллаген в течение 14С экстрагировали и очищали с использованием модифицированного метода Лонгина с ультрафильтрацией (желатин>30 кДа; (85)). Если выход коллагена был низким, а аминокислоты плохо сохранялись, мы использовали модифицированный процесс XAD (аминокислоты XAD; (86)). Для обеспечения качества мы измеряли концентрацию углерода и азота и соотношение C/N во всех извлеченных и очищенных образцах коллагена/аминокислот с помощью элементного анализатора Costech (ECS 4010). Мы оценивали качество по % выходу сырой желатины, %C, %N и соотношению C/N до AMS 14C-датирование. Соотношение C/N для всех образцов, непосредственно подвергшихся радиоуглеродному анализу, составляло от 2,9 до 3,6, что указывает на отличную сохранность (87). Образцы коллагена/аминокислот (~2,1 мг) затем сжигались в течение 3 часов при температуре 900°C в герметичных кварцевых трубках с проволоками из CuO и Ag. Образец CO2 был восстановлен до графита при 550°C с использованием H2 и катализатора Fe, при этом реакционная вода удалялась с помощью Mg(ClO4)2 (88). Образцы графита были спрессованы в мишени в алюминиевых лодочках и помещены на мишенное колесо вместе со стандартами OX-1 (щавелевая кислота), вторичными костями известного возраста и плейстоценовым китовым образцом, не содержащим 14C. Все измерения 14C проводились на модифицированном компактном спектрометре National Electronics Corporation с ускорителем на 0,5 МэВ (NEC 1.5SDH-1). Возраст 14C был скорректирован с учётом зависящей от массы фракционирования с измеренными значениями δ13C (89) и сравнен с образцами плейстоценовых китовых костей (фон, 48 000 14C лет назад), костей бизонов позднего голоцена (~1850 14C лет назад), коровьих костей конца 1800-х годов и стандартов щавелевой кислоты OX-2. Все калиброванные значения возраста 14C были рассчитаны с помощью OxCal версии 4.3 (Рэмси и Ли, 2013) с использованием кривой IntCal13 для северного полушария (90), и мы приводим 95-процентные доверительные интервалы (диапазоны 2-сигма).
Анализ основных компонентов (PCA)
Мы провели метод главных компонент с помощью пакета smartpca EIGENSOFT 7.2.1 (36). Мы использовали параметры по умолчанию и добавили две опции (lsqproject:YES и numoutlieriter:0), чтобы спроецировать древних особей в пространство главных компонент. Мы использовали два базовых набора данных для построения проекции: первый основан на генотипах 1340 современных евразийцев, генотипированных с помощью массива Affymetrix Human Origins, а второй — на подмножестве из 991 современного западноевразийца (7, 32, 33). Эти проекции неоднократно показаны в (13) и используются в онлайн-визуализаторе данных. Мы также вычислили FST между группами, используя параметры inbreed:YES и fstonly:YES. Мы ограничили этот анализ набором данных, полученным путём объединения наших данных о древней ДНК с данными о современной ДНК из массива Human Origins и ограничением до 597 573 однонуклеотидных полиморфизмов. Мы рассматривали позиции, для которых у нас не было данных о последовательности, как отсутствующие генотипы.
Кластеризация ПРИМЕСЕЙ
Используя PLINK2 (91), мы сначала сократили наш набор данных с помощью опции –geno 0.7, чтобы убедиться, что мы проводим анализ только в тех местах, где по крайней мере 70% индивидуумов имеют хотя бы одну последовательность. В результате получилось 892 613 однонуклеотидных полиморфизмов. Индивидуумам, не имеющим данных по конкретным однонуклеотидным полиморфизмам, были присвоены отсутствующие данные в этих местах. Мы запустили ADMIXTURE (37) с 10 репликами, выбрав реплику с наибольшей вероятностью. Мы приводим результаты для K=5 в (13), поскольку обнаружили, что это обеспечивает хорошее разрешение для определения источников происхождения древних людей до эпохи меди.
f-статистика
Мы использовали пакеты qp3pop и qpDstat в ADMIXTOOLS для вычисления f3-статистики и f4-статистики. Мы использовали параметр inbreed:YES для вычисления f3-статистики в качестве теста на смешение с древней популяцией в качестве цели и со всеми древними геномами в качестве источников. Используя параметр f4Mode:YES в qpDstat, мы также вычислили два набора статистики f4-симметрии, чтобы оценить, соответствуют ли пары популяций формированию клады по отношению к контрольной популяции. Первый набор — это статистика «Сравнение двух популяций», в которой мы сравниваем все возможные пары древних групп (популяции Test) с группой популяций, включающей различные популяции до медного века и более распространённые генетические вариации. Таким образом, мы вычисляем статистику в виде f4(Тест 1, Тест 2; Домедный век, Мбути). Вторая — это статистика «сходства с Домедным веком», которая сравнивает каждую древнюю группу со всеми возможными парами популяций Домедного века, используя статистику в виде f4(Домедный век 1, Домедный век 2; Тест, Мбути).
История моделирования примесей
Мы использовали qpAdm (33) в программном пакете ADMIXTOOLS для оценки пропорций происхождения в тестовой популяции, полученной в результате смешения N «эталонных» популяций, используя (но не моделируя явно) общий генетический дрейф с набором «внешних» популяций. Мы установили параметр details:YES, который сообщает о нормально распределённом Z-показателе соответствия модели (рассчитанном с помощью метода «блок-ножниц»).
Иерархическое моделирование.
Для каждой группы на предполагаемой клине мы использовали qpAdm, чтобы получить оценки доли происхождения от предполагаемых исходных популяций, а также ковариационную матрицу для разных групп. Мы совместно моделировали эти оценки с помощью двумерной нормальной модели (принуждая три доли в сумме составлять 100%) и оценивали среднее значение и ковариацию двух параметров с помощью метода максимального правдоподобия. С помощью полученной матрицы мы проверили, можно ли смоделировать клин с помощью смеси двух основных исходных популяций. Во-первых, мы проверили, соответствует ли ковариационная матрица единичному значению, что означает, что знание доли предков, относящихся к одному из компонентов смешения, позволяет полностью предсказать долю предков, относящихся к двум другим компонентам, как и ожидалось для двухкомпонентного смешения. Во-вторых, если бы нам удалось установить, что это так, мы бы изучили разницу между ожидаемыми и наблюдаемыми соотношениями долей предков анализируемых групп в рамках этой порождающей модели, подбирая все группы одновременно. В результате несколько групп отклонились от ожидаемых значений.
Метод датирования событий смешения.
Мы реализовали алгоритм DATES, который использует паттерны ковариации происхождения, которые можно измерить у одного человека (в отличие от LD-аддитивности, для которой требуется несколько человек). Подробная информация о подходе и результаты моделирования, демонстрирующие его эффективность как для современных, так и для древних данных, представлены в (13). Программное обеспечение, реализующее DATES, доступно по адресу https://zenodo.org/record/3263997#.XRnebJNKj6A (DOI: 10.5281/zenodo.3263997).
Great ?V I should certainly pronounce, impressed with your site. I had no trouble navigating through all the tabs as well as related information ended up being truly easy to do to access. I recently found what I hoped for before you know it in the least. Reasonably unusual. Is likely to appreciate it for those who add forums or anything, website theme . a tones way for your client to communicate. Nice task..