Механизм, который управляет режимом активности мозга.
Таламус человека на МРТ-снимке, отмечен стрелкойАвтор: Brian_chrischan CC BY-SA 3.0 Источник: commons.wikimedia.org
Долгое время основное внимание было приковано к коре головного мозга — его знаменитому серому веществу. Но последние исследования указывают на другую, более глубокую и центрально расположенную структуру — таламус.
Таламус — это не пассивный передатчик сенсорных сигналов. Это активная система управления, которая играет решающую роль и в поддержании общего состояния бодрствования, и в формировании конкретного содержания нашего восприятия. Разберем, как именно он это делает.
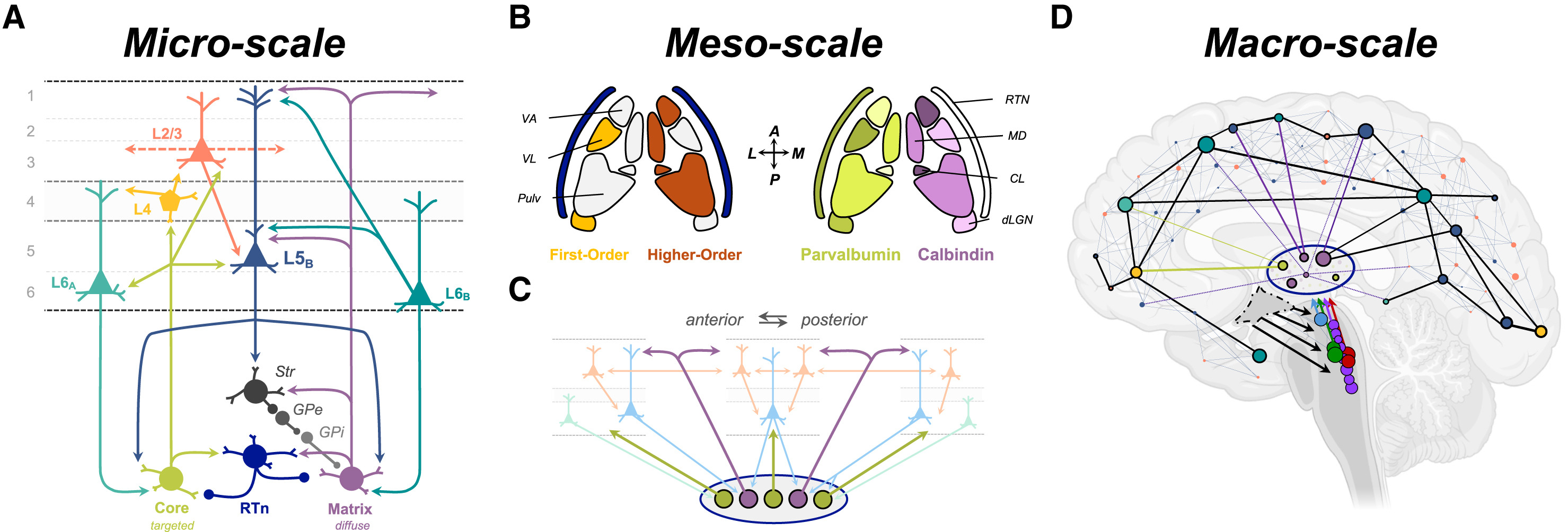
Микро-, мезо- и макроуровневая организация таламокортикальной системы (A) На микроуровне различные типы возбуждающих клеток в таламусе (ядерные [оливковый цвет] и матричные [сливовый цвет]) проецируются в различные слои (пронумерованные 1-6) коры головного мозга, где они контактируют со специфическими популяциями клеточных типов, которые оказывают различное влияние на кортикальные вычисления. Ядерные клетки посылают направленные проекции к звездчатым клеткам (L4, светло-оранжевый) в средних (зернистых) слоях, которые затем иннервируют пирамидальные нейроны в поверхностных (L2/3, мандариновый) и глубоких (L6A, бирюзовый) слоях, которые, в свою очередь, проецируются либо обратно в таламус (L6A), либо в пределах коры головного мозга (L2/3). В отличие от них, матричные клетки иннервируют поверхностные (L1) и глубокие (L5) слои, особенно нацеливаясь на крупные пирамидальные нейроны пирамидного тракта (PT-типа) с толстыми пучковыми дендритами (ttL5B, синий), которые являются единственным исходящим путем коры за пределы среднего мозга. И клетки ttL5B, и матричные клетки также тесно связаны с орексин-чувствительными пирамидальными нейронами слоя 6 (L6B, бирюзовый). Также существует обширный тормозной контроль над спайковой активностью таламуса: например, через ретикулярное ядро таламуса (RTn, темно-синий), которое возбуждается проекционными нейронами таламуса и кортикальными входами в таламус (не показано), и через внутренний бледный шар (GPi), который управляется входами от ttL5B к наружному бледному шару (GPe, средне-серый). NB: это идеализированная схема, предназначенная для общего обзора основных контуров, поэтому на ней показаны не все известные связи. (B) Различные ядра таламуса можно охарактеризовать в соответствии с их принадлежностью к (слева) первому порядку (светло-оранжевый) или высшему порядку (коричневый), в зависимости от того, поступают ли управляющие (драйверные) входы из подкорковых структур или из коры головного мозга соответственно; а также (справа) по экспрессии парвальбумина (в ядерных клетках) и кальбиндина (в матричных клетках). Обратите внимание, что отдельные ядра содержат смесь обоих типов клеток (что приблизительно показано относительной интенсивностью цвета; распределение внутри отдельных ядер не изображено). (C) На мезоуровне предполагается, что таламус контролирует локальную (через ядерные клетки) и дальнодействующую (через матричные клетки) возбудимость распределенных областей коры головного мозга посредством проекций, которые усиливают локальный или распределенный резонанс в кортико-таламических петлях по всей мантии коры. (D) На макроуровне таламус тесно взаимосвязан со всей мантией коры посредством аксональных связей, исходящих как из коры головного мозга, так и из таламуса, которые вместе образуют распределенную сеть, формирующую сознательный опыт. Особую важность имеет наличие в одной и той же системе как конвергентной (несколько кортикальных областей проецируются в одну область таламуса), так и дивергентной (одна область таламуса проецируется в несколько кортикальных областей) архитектуры. Гипоталамус (пунктирная линия) проецируется на множество узлов в восходящей системе возбуждения, где специализированные клетки высвобождают различные нейромодулирующие нейрохимические вещества (дофамин, светло-голубой; ацетилхолин, зеленый; норадреналин, темно-красный; серотонин, фиолетовый). Условные обозначения: CL, центральное латеральное ядро; dLGN, дорсальное латеральное коленчатое ядро; MD, медиодорсальное ядро; Pulv, подушка таламуса; RTN, ретикулярное ядро таламуса; VA, вентральное переднее ядро; VL, вентральное латеральное ядро.Автор: Whyte, Christopher J. et al. Neuron, Volume 112 Источник: www.cell.com
Два типа нейронов для двух задач
Все обусловлено двумя разными типами нейронов таламуса, которые работают в тесной связке с корой головного мозга. Разделяют их условно на «матричные» и «ядерные».
Матричные нейроны отвечают за состояние. Они обладают широкими, диффузными связями, охватывая большие участки коры. Их задача — создать общий фон, подготовить «сцену». Именно они переводят мозг в режим бодрствования, повышая общую возбудимость коры и делая ее готовой к приему информации.
Ядерные нейроны отвечают за содержание. Их связи гораздо более точные и сфокусированные, работающие с конкретными функциональными зонами коры. Именно они обеспечивают обработку специфических сигналов — того, что мы видим или слышим в данный момент.
Но как именно они это делают?
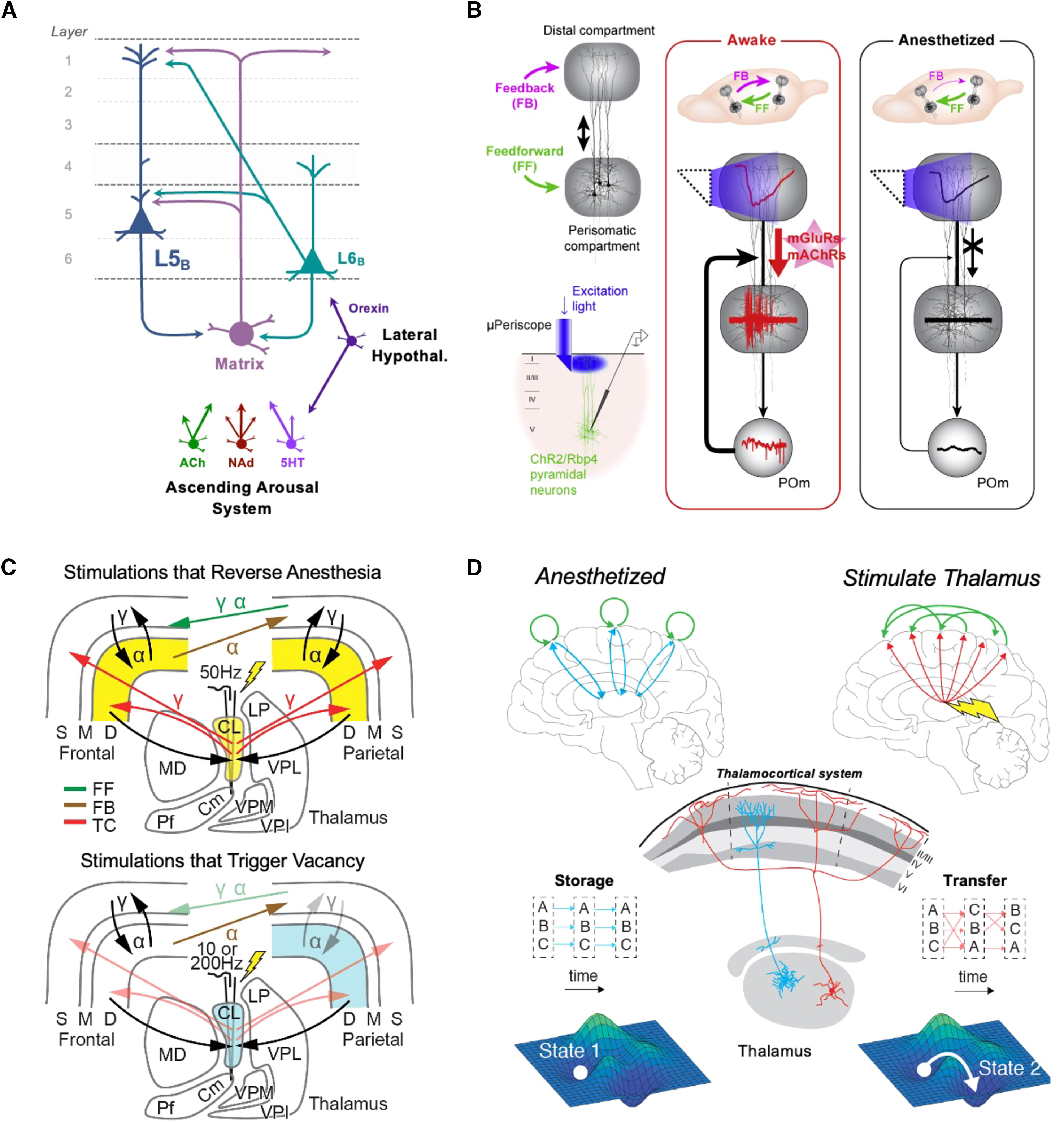
Взаимодействие матричных входов таламуса с корой головного мозга на разных масштабах для изменения состояния сознания (A) Таламокортикальные контуры, вовлеченные в состояние сознания. Матричные клетки таламуса (фиолетовые) являются важными мишенями ретикулярной активирующей системы. Они взаимно связаны с нейронами ttL5B (темно-синие) и идеально расположены для модуляции их возбудимости. Нейроны L6B, управляемые орексином, дополнительно модулируют эти таламокортикальные взаимодействия. (B) Во время бодрствования матричные входы таламуса возбуждают пирамидальные нейроны L5 через метаботропные глутаматергические входы к косым дендритам, позволяя кальциевым спайкам в апикальных дендритах влиять на соматическую активность. Под общей анестезией, в отсутствие таламического возбуждения, пирамидальные нейроны L5 не способны распространять сигналы от дендритов к соме, что способствует функциональному разобщению областей мозга. Рисунок адаптирован с разрешения Suzuki and Larkum.⁸⁰ (C) Минимально достаточный механизм, посредством которого DBS центрального латерального ядра (CL) таламуса восстанавливает или нарушает сознание. Верхняя часть: таламокортикальные (TC), прямые (FF) и обратные (FB) связи, наиболее измененные стимуляцией таламуса, которая успешно обращала общую анестезию (пропофол и изофлуран) у макак. Высокочастотная (50 Гц) DBS таламуса, одновременно подаваемая через 16 контактов линейного электрода (с шагом 200 мкм), центрированного в CL, имитирует паттерны активности бодрствующего состояния (желтый цвет), характерные для таламокортикальных эфферентов. Это увеличивает частоту разрядов в глубоких слоях коры (желтый цвет), что восстанавливает внутриколоночные, прямые и обратные связи на альфа- и гамма-частотах, несмотря на тормозное давление анестезии. Адаптировано с разрешения Redinbaugh et al.⁸¹ Нижняя часть: низко- (10 Гц) и очень высокочастотная (200 Гц) DBS, центрированная в CL, у бодрствующих макак вместо этого вызывает «пустое» состояние, схожее с симптомами абсансной эпилепсии. Низкая активность в таламусе и глубоких слоях коры (синий цвет) приводит к нарушению связей в теменных внутриколоночных контурах и разрыву прямых связей (светлая штриховка). На основе результатов Redinbaugh et al.⁸² (D) Крупномасштабная кортико-таламическая модель нейронных масс (включающая матричные и ядерные клетки таламуса), которая воспроизвела восстановление сознания при стимуляции таламуса после пропофоловой анестезии у макак. Симуляция стимуляции матричных, но не ядерных, клеток повышала кортикальную возбудимость, обеспечивая передачу информации между областями и более сложную, менее стереотипную динамику. На макроуровне это отразилось в «уплощении» общемозгового энергетического ландшафта, что типично для состояния бодрствования. Рисунок воспроизведен с разрешения Müller et al.⁸³Автор: Whyte, Christopher J. et al. Neuron, Volume 112 Источник: www.cell.com
Как включить мозг: механизм бодрствования
Между глубокими слоями коры и таламусом существует постоянная петля обратной связи: глубокие слои коры посылают сигналы в таламус, а таламус отвечает им обратно. Ключевую роль в этой петле играют матричные нейроны.
Когда вы бодрствуете, эта петля активна. Сигналы курсируют, поддерживая кору в «горячем», готовом к работе состоянии. Что происходит под общей анестезией? Анестетики не выключают кору напрямую. Они разрывают именно эту связь между корой и таламусом. Петля замолкает, и сознание гаснет.
Здесь кроется самое интересное. Эксперименты с глубокой стимуляцией мозга (DBS) показали, что если подать электрический импульс на определенные участки таламуса у пациента под наркозом, эту петлю можно запустить принудительно. Результат? Человек начинает проявлять признаки сознания, даже пока анестетик все еще действует.
Таламус, а точнее его матричные нейроны, не просто реагирует на сигналы пробуждения от ствола мозга. Он является активным участником, который «включает» кору и делает сознательный опыт возможным.
Хорошо, «свет» включили. Но что именно мы увидим на «экране»?
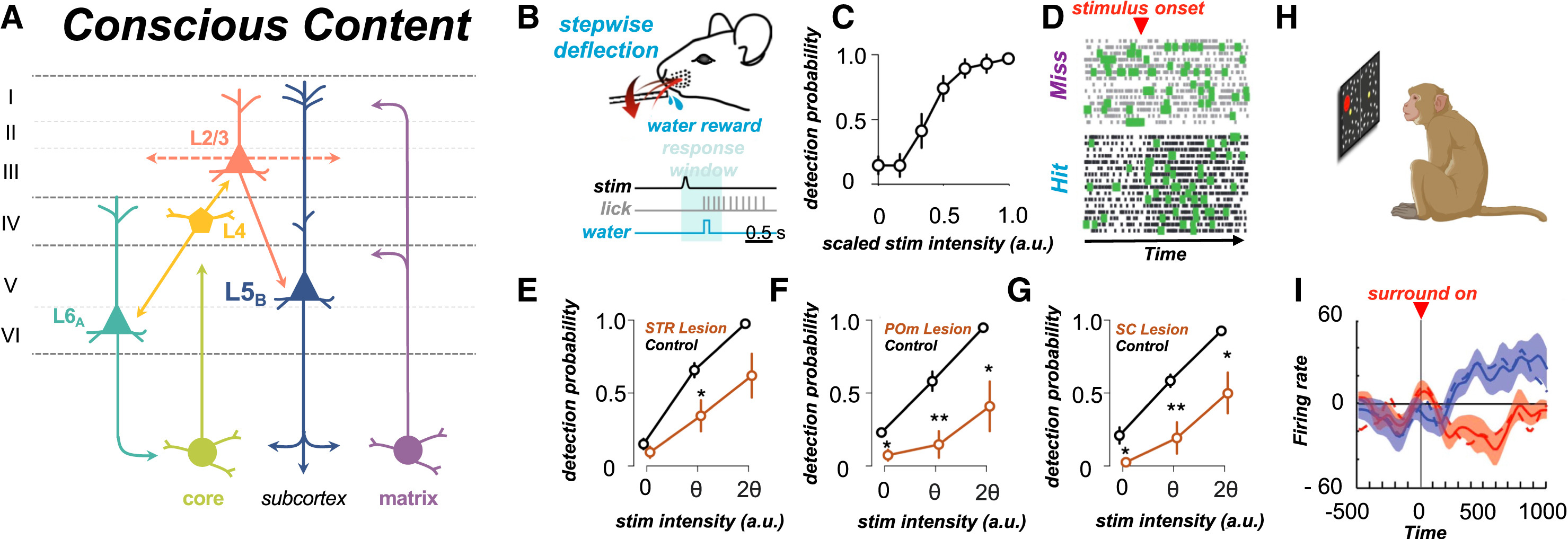
Вклад таламуса в содержание сознания (A) Гипотетический таламокортикальный контур, поддерживающий содержание сознания. Матричные клетки таламуса (фиолетовые) проецируются к апикальным дендритам клеток ttL5B (темно-синие) и пирамидальных клеток L2/3 (оранжевые), а также к косым дендритам клеток ttL5B в слое L5A. Проекции к косым дендритам деполяризуют ствол клетки через действие метаботропных глутаматергических (и холинергических) рецепторов, что, как было предположено и показано, облегчает связь между апикальным и соматическим компартментами, позволяя натриевым спайкам распространяться в обратном направлении от сомы к апикальным дендритам, способствуя генерации кальциевых спайков, и наоборот. И матричные, и ядерные (светло-зеленые) клетки таламуса получают управляющие (драйверные) входы от клеток ttL5B. Ядерные клетки посылают управляющие проекции к IT-клеткам L4 и пирамидальным клеткам L2/3 (не показано). IT-клетки L4 затем формируют замкнутый возбуждающий контур с ядерными клетками через нейроны L6A (голубые). Аналогичным образом, возбуждающий выход из клеток L4 нацелен на базальные дендриты клеток ttL5B через слой L2/3. (B) Иллюстрация парадигмы порогового обнаружения, использованной в исследованиях Takahashi и его коллег.¹⁶,¹⁴⁵ (C) Психометрическая функция, показывающая вероятность ответа (среднее +- SD) как функцию (масштабированной) интенсивности стимула у мышей во время сеансов записи. (D) Пример растровых графиков для сомы клеток ttL5B в пробах с попаданием (hit) и промахом (miss). Редкие вспышки (зеленый цвет) происходят в обоих условиях, но в пробах с попаданием они синхронизированы по времени с началом стимула. (E-G) Вероятность обнаружения (среднее +- SEM) в парадигме порогового обнаружения как функция интенсивности стимула при сравнении контрольного условия с (E) торможением стриатума, (F) торможением POm или (G) торможением верхнего двухолмия, из работы Takahashi et al.¹⁶ (H) Обобщенная парадигма подавления вспышкой, использованная Wilke et al.¹⁴⁶ Обезьяны фиксировали взгляд на центральной точке в течение 1,5 с, после чего на периферии зрения появлялся целевой стимул (красный диск). Через 2 с после появления целевого стимула появлялся окружающий узор, делая целевой стимул невидимым с вероятностью, пропорциональной плотности узора. Обезьян обучали сообщать об исчезновении стимула нажатием на рычаг. (I) Средняя частота разрядов (+-SEM) нейронов в дорсальной и вентральной подушке таламуса, активность которых подавлялась при исчезновении стимула (красный) или активировалась при исчезновении стимула (синий). Пунктирные линии показывают реакции подушки таламуса на физическое удаление стимула, в то время как сплошные линии показывают реакции подушки на воспринимаемое исчезновение. (B)-(I) были воссозданы с разрешения Takahashi et al.¹⁶,¹⁴⁵ и Wilke et al.¹⁴⁶Автор: Whyte, Christopher J. et al. Neuron, Volume 112 Источник: www.cell.com
Что именно мы осознаем? Таламус за пультом
Здесь в игру вступают ядерные нейроны. Они отвечают за то, какая именно информация станет частью сознательного опыта.
Возьмем простой пример: попытка расслышать тихий шорох в ночной тишине. Сигнал от ушей идет в мозг, но станет ли он осознанным? Оказывается, это во многом зависит от активности в петле между корой и ядерными нейронами таламуса.
Исследования на животных показали: когда слабый сенсорный стимул (например, легкое касание уса) осознается, в этой петле возникает специфическая вспышка активности. Если вспышки нет, стимул остается «за кадром», хоть органы чувств его и зафиксировали. Таламус буквально управляет порогом восприятия.
Более того, он обеспечивает стабильность нашего мира. Каждый раз, когда ваши глаза совершают быстрое движение (саккаду), изображение на сетчатке смазывается и смещается. Почему же мир не дергается перед глазами несколько раз в секунду? Потому что таламус получает копию команды на движение глаз и заранее «готовит» кору, обеспечивая бесшовное восприятие. Он сглаживает переходы между кадрами.
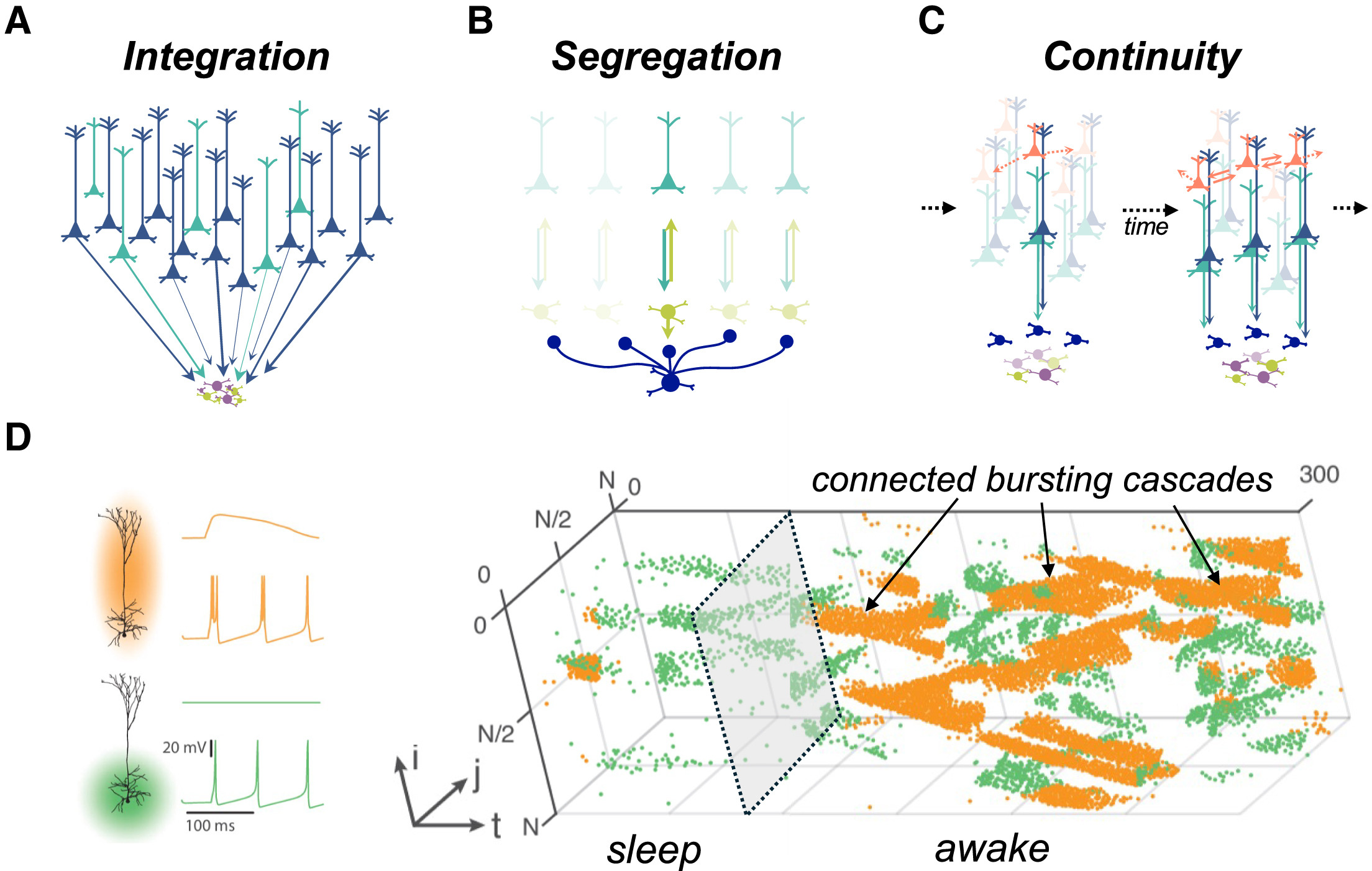
Вклад таламуса в характер содержания сознания (A-C) Ограничения, налагаемые на текущую динамику организацией таламуса, могут помочь объяснить ключевые особенности обработки сознательной информации, такие как (A) интеграция, которая, как мы утверждаем, возникает в результате снижения размерности, налагаемого сходящимися входами на меньшее количество клеток в таламусе (нижний ряд) по сравнению с корой головного мозга (верхний ряд); (B) сегрегация, которая является естественным побочным продуктом процесса, подобного разделяющей нормализации, возникающего из-за зависимого от активности привлечения тормозного ретикулярного ядра таламуса (темно-синий); и (C) непрерывность, при которой сочетание управляющей (драйверной) и модулирующей ролей в таламусе налагает «эффект Матфея» на клеточные популяции, что дает импульс активности нейронам, связанным с текущим доминирующим таламокортикальным ансамблем. Цвета клеток в (A)-(C) соответствуют цветам на Рисунках 1, 2 и 3. (D) Была создана спайковая нейронная модель для имитации ключевой особенности клеток ttL5B, а именно, их перехода от режима регулярных спайков (зеленый; вход только к базальным дендритам) к режиму вспышек (оранжевый; одновременная активация апикальных и базальных дендритов), когда он управляется матричными входами таламуса. После настройки модели по электрофизиологическим данным, полученным от макак под анестезией/бодрствующих и естественно спящих, авторы показали, что область в пространстве параметров модели, которая лучше всего соответствует сну, была связана с преимущественно регулярной спайковой активностью (зеленые точки), тогда как в режиме бодрствования (следуя за серой пунктирной моделью), модельные нейроны ttL5B (встроенные в кортикальный слой размером N = 100 x 100) были способны переходить в режим вспышек, который, в свою очередь, запускал вспышки в синаптически связанных нейронах, приводя к устойчивой активности, которая могла когерентно распространяться по кортикальному слою — процесс, названный «связанные каскады вспышек». (D) было адаптировано из Munn et al.¹⁴⁸ с разрешения. Обратите внимание, что эти механизмы предназначены лишь для того, чтобы передать правдоподобные, но не исчерпывающие способы реализации каждой из этих особенностей.Автор: Whyte, Christopher J. et al. Neuron, Volume 112 Источник: www.cell.com
Три кита нашего опыта: как таламус создает целостную реальность
Но роль таламуса еще глубже. Сама структура его связей с корой объясняет три фундаментальных свойства нашего сознания: его целостность, разделенность и непрерывность.
Интеграция (Целостность). Почему яблоко воспринимается как единый объект — красное, круглое, гладкое, — а не как набор отдельных признаков? В коре мозга работают миллиарды нейронов, обрабатывающих эти признаки в разных зонах. А в таламусе нейронов на порядки меньше. Когда вся эта информация из коры сходится в таламусе, она неизбежно «сжимается». Этот процесс, известный как снижение размерности, заставляет мозг объединять разрозненные потоки данных в единый, интегрированный образ.
Сегрегация (Разделенность). Как у вас получается сосредоточиться на этой статье, игнорируя шум за окном? Вокруг таламуса есть тормозная оболочка — ретикулярное ядро (TRN). Когда одна таламо-кортикальная петля (обрабатывающая текст) становится активной, она возбуждает TRN, которое, в свою очередь, подавляет соседние, конкурирующие петли (обрабатывающие шум). Это механизм, который позволяет одному потоку информации доминировать в сознании, отсекая все лишнее.
Непрерывность. Почему сознание ощущается как плавный, непрерывный поток, а не набор случайных вспышек? Активная в данный момент таламо-кортикальная петля имеет преимущество. Ей проще оставаться активной или повлиять на то, какая петля активируется следующей. Возникает своего рода инерция, которая и создает ощущение плавного, непрерывного течения опыта.
Не просто реле, а центральный процессор
Таким образом, таламус предстает не пассивным посредником, а динамической системой управления. Он использует разные типы нейронов для двух ключевых задач: поддержания общего состояния бодрствования и формирования конкретного содержания нашего опыта.
Он сжимает информацию, чтобы создать целостный мир, отсекает лишнее, чтобы мы могли сфокусироваться, и обеспечивает плавность нашего восприятия.
Фундаментальный вывод заключается в том, что сознание — это не продукт одной лишь коры головного мозга. Это результат сложного и непрерывного взаимодействия между корой и таламусом, где каждая из структур выполняет свою незаменимую роль.